Consumption of a Specially-Formulated Mixture of Essential Amino Acids Promotes Gain in Whole-Body Protein to a Greater Extent than a Complete Meal Replacement in Older Women with Heart Failure


Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
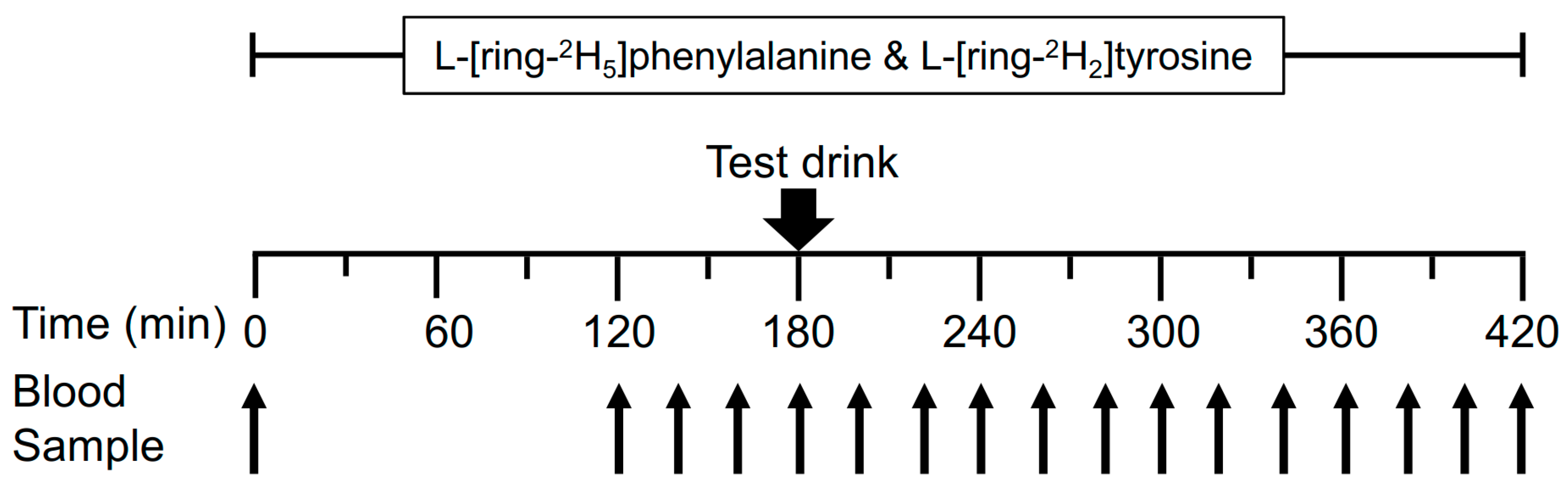
2.2. Experimental Design
2.3. Test Drinks
2.4. Stable Isotope Tracer Infusion Protocol
2.5. Analytic Methods
2.6. Calculations of Protein Kinetics
2.7. Statistical Analysis
3. Results
3.1. Baseline Characteristics
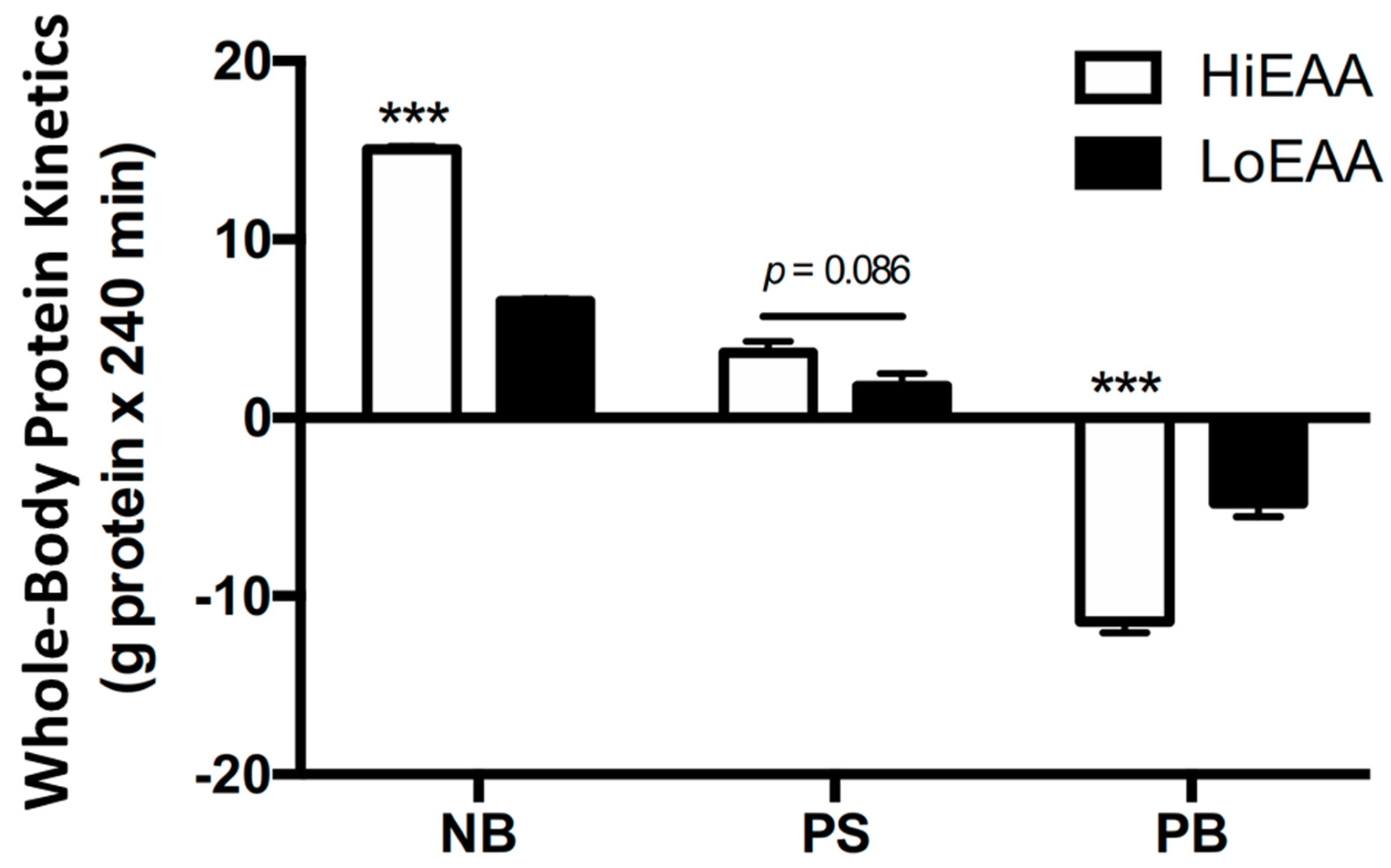
3.2. Whole-Body Protein Kinetics
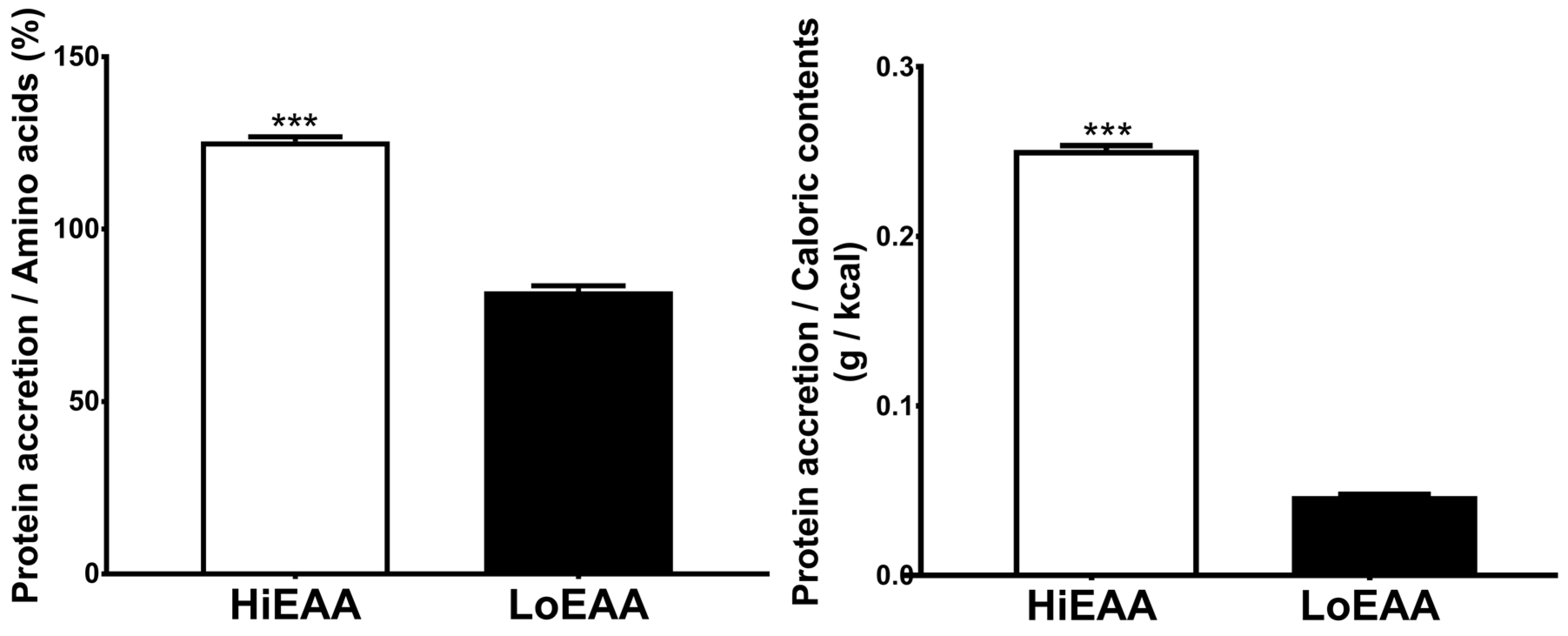
3.3. Protein Accretion Efficiency
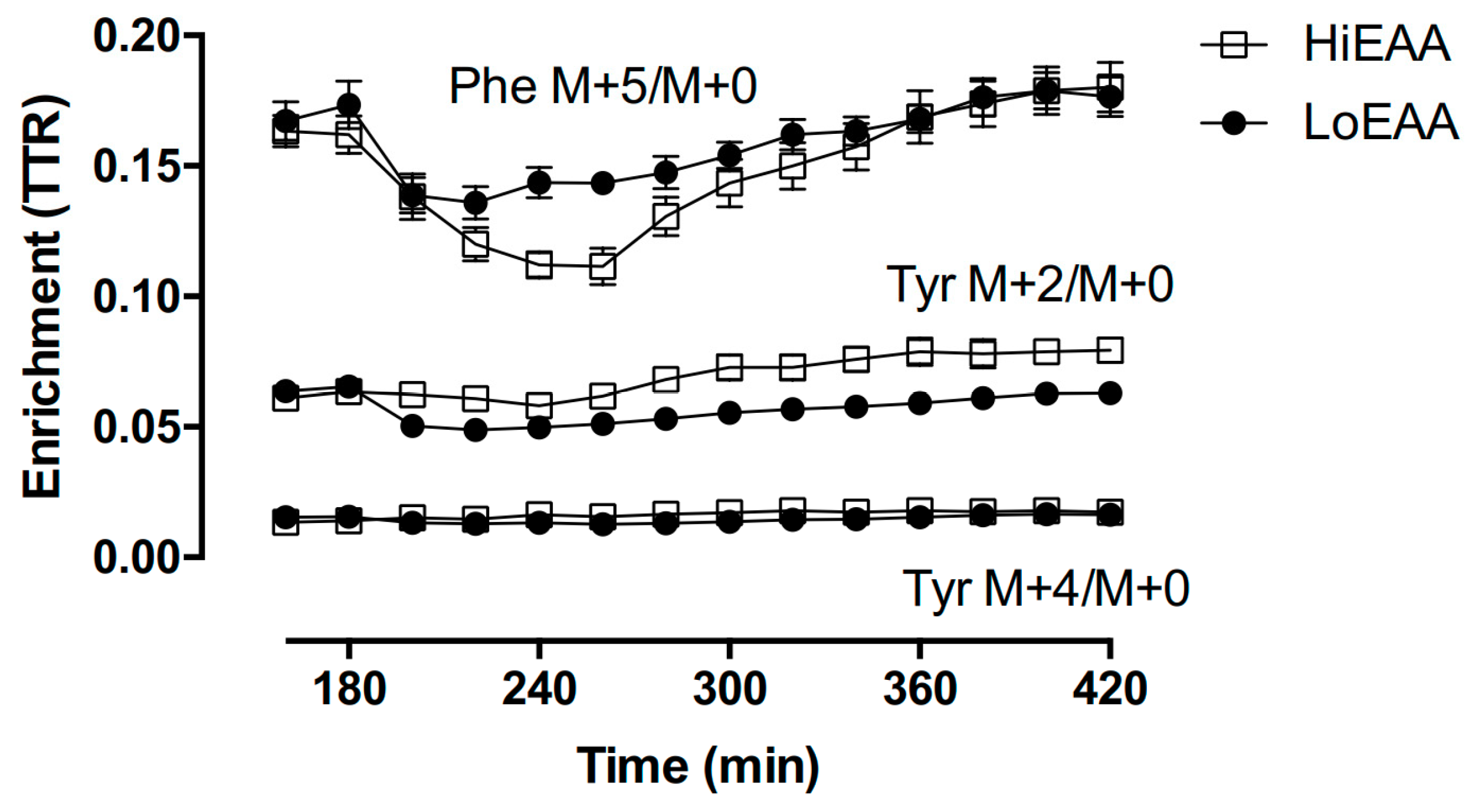
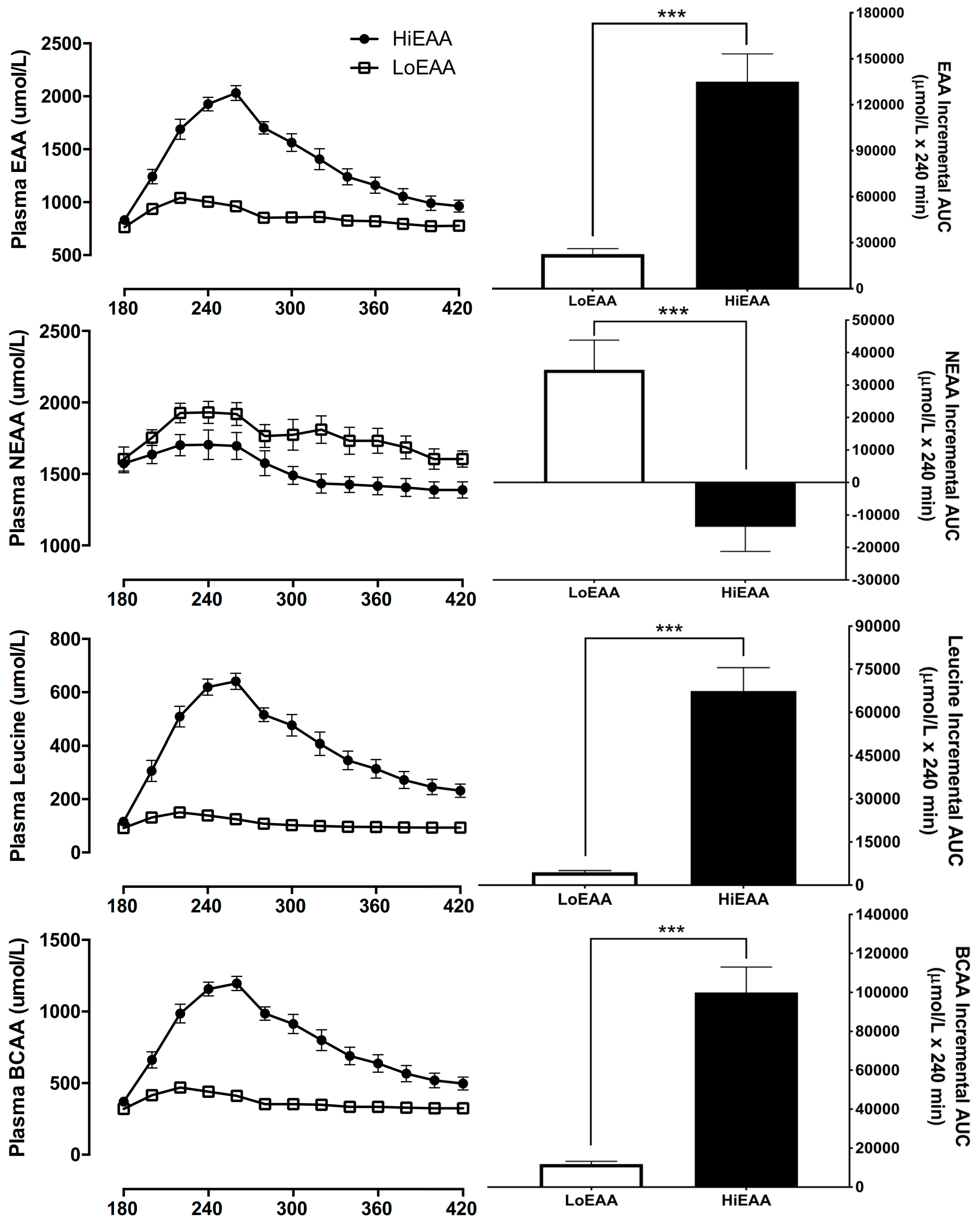
3.4. Plasma Amino Acid Responses
4. Discussion
5. Conclusion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- McMurray, J.J.; Pfeffer, M.A. Heart failure. Lancet 2005, 365, 1877–1889. [Google Scholar] [CrossRef]
- The National Clinical Guideline Centre. Chronic Heart Failure; Royal College of Physicians: London, UK, 2010. [Google Scholar]
- Azhar, G.; Wei, J.Y. New approaches to treating cardiac cachexia in the older patient. Curr. Cardiovasc. Risk Rep. 2013, 7, 480–484. [Google Scholar] [CrossRef]
- Bekfani, T.; Pellicori, P.; Morris, D.A.; Ebner, N.; Valentova, M.; Steinbeck, L.; Wachter, R.; Elsner, S.; Sliziuk, V.; Schefold, J.C.; et al. Sarcopenia in patients with heart failure with preserved ejection fraction: Impact on muscle strength, exercise capacity and quality of life. Int. J. Cardiol. 2016, 222, 41–46. [Google Scholar] [CrossRef] [Green Version]
- Anker, S.D.; Sharma, R. The syndrome of cardiac cachexia. Int. J. Cardiol. 2002, 85, 51–66. [Google Scholar] [CrossRef]
- Upadhya, B.; Haykowsky, M.J.; Eggebeen, J.; Kitzman, D.W. Exercise intolerance in heart failure with preserved ejection fraction: More than a heart problem. J. Geriatr. Cardiol. 2015, 12, 294–304. [Google Scholar] [CrossRef]
- Wang, W.; Jiang, T.; Li, C.; Chen, J.; Cao, K.; Qi, L.-W.; Li, P.; Zhu, W.; Zhu, B.; Chen, Y. Will testosterone replacement therapy become a new treatment of chronic heart failure? A review based on 8 clinical trials. J. Thorac. Dis. 2016, 8, E269–E277. [Google Scholar] [CrossRef] [Green Version]
- Pasini, E.; Aquilani, R.; Gheorghiade, M.; Dioguardi, F.S. Malnutrition, muscle wasting and cachexia in chronic heart failure: The nutritional approach. Ital. Heart J. 2003, 4, 232–235. [Google Scholar]
- Hibbert, J.M.; Broemeling, L.D.; Isenberg, J.N.; Wove, R.R. Determinants of free-living energy expenditure in normal weight and obese women measured by doubly labeled water. Obes. Res. 1994, 2, 44–53. [Google Scholar] [CrossRef]
- Bouchard, D.R.; Dionne, I.J.; Brochu, M. Sarcopenic/obesity and physical capacity in older men and women: Data from the nutrition as a determinant of successful aging (NuAge)—The Quebec longitudinal study. Obesity 2009, 17, 2082–2088. [Google Scholar] [CrossRef]
- Toth, M.J.; Matthews, D.E. Whole-body protein metabolism in chronic heart failure: Relationship to anabolic and catabolic hormones. J. Parenter. Enter. Nutr. 2006, 30, 194–201. [Google Scholar] [CrossRef]
- Morrison, W.L.; Gibson, J.N.A.; Rennie, M.J. Skeletal muscle and whole body protein turnover in cardiac cachexia: Influence of branched-chain amino acid administration. Eur. J. Clin. Investig. 1988, 18, 648–654. [Google Scholar] [CrossRef]
- Van Dijk, D.P.; van de Poll, M.C.; Moses, A.G.; Preston, T.; Olde Damink, S.W.; Rensen, S.S.; Deutz, N.E.; Soeters, P.B.; Ross, J.A.; Fearon, K.C.; et al. Effects of oral meal feeding on whole body protein breakdown and protein synthesis in cachectic pancreatic cancer patients. J. Cachexia Sarcopenia Muscle 2015, 6, 212–221. [Google Scholar] [CrossRef]
- Katsanos, C.S.; Kobayashi, H.; Sheffield-Moore, M.; Aarsland, A.; Wolfe, R.R. Aging is associated with diminished accretion of muscle proteins after the ingestion of a small bolus of essential amino acids. Am. J. Clin. Nutr. 2005, 82, 1065–1073. [Google Scholar] [CrossRef] [Green Version]
- Katsanos, C.S.; Kobayashi, H.; Sheffield-Moore, M.; Aarsland, A.; Wolfe, R.R. A high proportion of leucine is required for optimal stimulation of the rate of muscle protein synthesis by essential amino acids in the elderly. Am. J. Physiol. Endocrinol. Metab. 2006, 291, E381–E387. [Google Scholar] [CrossRef] [Green Version]
- Børsheim, E.; Bui, Q.-U.T.; Tissier, S.; Kobayashi, H.; Ferrando, A.A.; Wolfe, R.R. Effect of amino acid supplementation on muscle mass, strength and physical function in elderly. Clin. Nutr. 2008, 27, 189–195. [Google Scholar] [CrossRef] [Green Version]
- Bukhari, S.S.I.; Phillips, B.E.; Wilkinson, D.J.; Limb, M.C.; Rankin, D.; Mitchell, W.K.; Kobayashi, H.; Greenhaff, P.L.; Smith, K.; Atherton, P.J. Intake of low-dose leucine-rich essential amino acids stimulates muscle anabolism equivalently to bolus whey protein in older women at rest and after exercise. Am. J. Physiol. Endocrinol. Metab. 2015, 308, E1056–E1065. [Google Scholar] [CrossRef]
- Børsheim, E.; Tipton, K.D.; Wolf, S.E.; Wolfe, R.R. Essential amino acids and muscle protein recovery from resistance exercise. Am. J. Physiol. Endocrinol. Metab. 2002, 283, E648–E657. [Google Scholar] [CrossRef] [Green Version]
- Paddon-Jones, D.; Sheffield-Moore, M.; Urban, R.J.; Aarsland, A.; Wolfe, R.R.; Ferrando, A.A. The catabolic effects of prolonged inactivity and acute hypercortisolemia are offset by dietary supplementation. J. Clin. Endocrinol. Metab. 2005, 90, 1453–1459. [Google Scholar] [CrossRef]
- Biolo, G.; Gastaldelli, A.; Zhang, X.J.; Wolfe, R.R. Protein synthesis and breakdown in skin and muscle: A leg model of amino acid kinetics. Am. J. Physiol. Endocrinol. Metab. 1994, 267, E467–E474. [Google Scholar] [CrossRef]
- Kim, I.-Y.; Schutzler, S.; Schrader, A.; Spencer, H.; Kortebein, P.; Deutz, N.E.P.; Wolfe, R.R.; Ferrando, A.A. Quantity of dietary protein intake, but not pattern of intake, affects net protein balance primarily through differences in protein synthesis in older adults. Am. J. Physiol. Endocrinol. Metab. 2015, 308, E21–E28. [Google Scholar] [CrossRef]
- Kim, I.-Y.; Schutzler, S.; Schrader, A.M.; Spencer, H.J.; Azhar, G.; Wolfe, R.R.; Ferrando, A.A. Protein intake distribution pattern does not affect anabolic response, lean body mass, muscle strength or function over 8 weeks in older adults: A randomized-controlled trial. Clin. Nutr. 2018, 37, 488–493. [Google Scholar] [CrossRef]
- Kim, I.-Y.; Shin, Y.-A.; Schutzler, S.E.; Azhar, G.; Wolfe, R.R.; Ferrando, A.A. Quality of meal protein determines anabolic response in older adults. Clin. Nutr. 2018, 37, 2076–2083. [Google Scholar] [CrossRef]
- Kim, I.-Y.; Schutzler, S.; Schrader, A.; Spencer, H.J.; Azhar, G.; Ferrando, A.A.; Wolfe, R.R. The anabolic response to a meal containing different amounts of protein is not limited by the maximal stimulation of protein synthesis in healthy young adults. Am. J. Physiol. Endocrinol. Metab. 2016, 310, E73–E80. [Google Scholar] [CrossRef] [Green Version]
- Wolfe, R.R.; Park, S.; Kim, I.-Y.; Starck, C.; Marquis, B.J.; Ferrando, A.A.; Moughan, P.J. Quantifying the contribution of dietary protein to whole body protein kinetics: Examination of the intrinsically-labeled proteins method. Am. J. Physiol. Endocrinol. Metab. 2019. [Google Scholar] [CrossRef]
- Rutherfurd, S.M.; Fanning, A.C.; Miller, B.J.; Moughan, P.J. Protein digestibility-corrected amino acid scores and digestible indispensable amino acid scores differentially describe protein quality in growing male rats. J. Nutr. 2015, 145, 372–379. [Google Scholar] [CrossRef]
- Wolfe, R.R.; Chinkes, D.L. Isotope Tracers in Metabolic Research: Principles and Practice of Kinetic Analysis; Wiley-Liss: New York, NY, USA, 2005; ISBN 9780471462095. [Google Scholar]
- Kim, I.-Y.; Deutz, N.E.P.; Wolfe, R.R. Update on maximal anabolic response to dietary protein. Clin. Nutr. 2018, 37, 411–418. [Google Scholar] [CrossRef]
- Paddon-Jones, D.; Sheffield-Moore, M.; Katsanos, C.S.; Zhang, X.-J.; Wolfe, R.R. Differential stimulation of muscle protein synthesis in elderly humans following isocaloric ingestion of amino acids or whey protein. Exp. Gerontol. 2006, 41, 215–219. [Google Scholar] [CrossRef]
- Deutz, N.E.; Wolfe, R.R. Is there a maximal anabolic response to protein intake with a meal? Clin. Nutr. 2013, 32, 309–313. [Google Scholar] [CrossRef]
- Bohé, J.; Low, A.; Wolfe, R.R.; Rennie, M.J. Human muscle protein synthesis is modulated by extracellular, not intramuscular amino acid availability: A dose-response study. J. Physiol. 2003, 552, 315–324. [Google Scholar] [CrossRef]
- Areta, J.L.; Burke, L.M.; Camera, D.M.; West, D.W.D.; Crawshay, S.; Moore, D.R.; Stellingwerff, T.; Phillips, S.M.; Hawley, J.A.; Coffey, V.G. Reduced resting skeletal muscle protein synthesis is rescued by resistance exercise and protein ingestion following short-term energy deficit. Am. J. Physiol. Endocrinol. Metab. 2014, 306, E989–E997. [Google Scholar] [CrossRef]
- Miller, A.L.; Tipton, K.D.; Chinkes, D.L.; Wolf, S.E.; Wolfe, R.R. Independent and combined effects of amino acids and glucose after resistance exercise. Med. Sci. Sports Exerc 2003, 35, 449–455. [Google Scholar] [CrossRef]
- Katsanos, C.S.; Aarsland, A.; Cree, M.G.; Wolfe, R.R. Muscle protein synthesis and balance responsiveness to essential amino acids ingestion in the presence of elevated plasma free fatty acid concentrations. J. Clin. Endocrinol. Metab. 2009, 94, 2984–2990. [Google Scholar] [CrossRef]
- Volpi, E.; Kobayashi, H.; Sheffield-Moore, M.; Mittendorfer, B.; Wolfe, R.R. Essential amino acids are primarily responsible for the amino acid stimulation of muscle protein anabolism in healthy elderly adults. Am. J. Clin. Nutr. 2003, 78, 250–258. [Google Scholar] [CrossRef]
- Anthony, J.C.; Anthony, T.G.; Kimball, S.R.; Vary, T.C.; Jefferson, L.S. Orally administered leucine stimulates protein synthesis in skeletal muscle of postabsorptive rats in association with increased eIF4F formation. J. Nutr. 2000, 130, 139–145. [Google Scholar] [CrossRef]
- Miller, S.; Chinkes, D.; MacLean, D.A.; Gore, D.; Wolfe, R.R. In vivo muscle amino acid transport involves two distinct processes. Am. J. Physiol. Endocrinol. Metab. 2004, 287, E136–E141. [Google Scholar] [CrossRef]
- Luiking, Y.C.; Ten Have, G.A.M.; Wolfe, R.R.; Deutz, N.E.P. Arginine de novo and nitric oxide production in disease states. Am. J. Physiol. Endocrinol. Metab. 2012, 303, E1177–E1189. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Chinkes, D.L.; Wolfe, R.R. The anabolic effect of arginine on proteins in skin wound and muscle is independent of nitric oxide production. Clin. Nutr. 2008, 27, 649–656. [Google Scholar] [CrossRef]
- Kim, I.-Y.; Schutzler, S.E.; Schrader, A.; Spencer, H.J.; Azhar, G.; Deutz, N.E.P.; Wolfe, R.R. Acute ingestion of citrulline stimulates nitric oxide synthesis but does not increase blood flow in healthy young and older adults with heart failure. Am. J. Physiol. Endocrinol. Metab. 2015, 309, E915–E924. [Google Scholar] [CrossRef] [Green Version]
- Udani, S.M.; Koyner, J.L. The effects of heart failure on renal function. Cardiol. Clin. 2010, 28, 453–465. [Google Scholar] [CrossRef]
- Jankowski, C.M.; Gozansky, W.S.; Van Pelt, R.E.; Schenkman, M.L.; Wolfe, P.; Schwartz, R.S.; Kohrt, W.M. Relative contributions of adiposity and muscularity to physical function in community-dwelling older adults. Obesity (Silver Spring) 2008, 16, 1039–1044. [Google Scholar] [CrossRef]
- Paddon-Jones, D.; Sheffield-Moore, M.; Aarsland, A.; Wolfe, R.R.; Ferrando, A.A. Exogenous amino acids stimulate human muscle anabolism without interfering with the response to mixed meal ingestion. Am. J. Physiol. Endocrinol. Metab. 2005, 288, E761–E767. [Google Scholar] [CrossRef] [Green Version]
- Dillon, E.L.; Sheffield-Moore, M.; Paddon-Jones, D.; Gilkison, C.; Sanford, A.P.; Casperson, S.L.; Jiang, J.; Chinkes, D.L.; Urban, R.J. Amino acid supplementation increases lean body mass, basal muscle protein synthesis, and insulin-like growth factor-I expression in older women. J. Clin. Endocrinol. Metab. 2009, 94, 1630–1637. [Google Scholar] [CrossRef]
- Ferrando, A.A.; Paddon-Jones, D.; Hays, N.P.; Kortebein, P.; Ronsen, O.; Williams, R.H.; McComb, A.; Symons, T.B.; Wolfe, R.R.; Evans, W. EAA supplementation to increase nitrogen intake improves muscle function during bed rest in the elderly. Clin. Nutr. 2010, 29, 18–23. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Age (year) | 80 ± 7 a |
| BMI (kg/m2) | 32.7 ± 5.9 |
| Body Weight (kg) | 81.9 ± 16 |
| Lean Body Mass (kg) | 41.0 ± 7.0 |
| Body Fat (%) | 45.5 ± 3.2 |
| Current Smoker (n, %) | 0 (0%) |
| Systolic BP (mm Hg) | 145 ± 17 |
| Diastolic BP (mm Hg) | 75 ± 5 |
| Heart rate (beats/min) | 73 ± 12 |
| MMSE b | 28 ± 2 |
| Medication (n, %) c | |
| ACE inhibitor | 4 (50%) |
| β-blocker | 3 (37.5%) |
| Diuretics | 6 (75%) |
| Angiotensin II receptor blockers | 0 (0%) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, I.-Y.; Park, S.; Smeets, E.T.H.C.; Schutzler, S.; Azhar, G.; Wei, J.Y.; Ferrando, A.A.; Wolfe, R.R. Consumption of a Specially-Formulated Mixture of Essential Amino Acids Promotes Gain in Whole-Body Protein to a Greater Extent than a Complete Meal Replacement in Older Women with Heart Failure. Nutrients 2019, 11, 1360. https://doi.org/10.3390/nu11061360
Kim I-Y, Park S, Smeets ETHC, Schutzler S, Azhar G, Wei JY, Ferrando AA, Wolfe RR. Consumption of a Specially-Formulated Mixture of Essential Amino Acids Promotes Gain in Whole-Body Protein to a Greater Extent than a Complete Meal Replacement in Older Women with Heart Failure. Nutrients. 2019; 11(6):1360. https://doi.org/10.3390/nu11061360
Chicago/Turabian StyleKim, Il-Young, Sanghee Park, Ellen T. H. C. Smeets, Scott Schutzler, Gohar Azhar, Jeanne Y. Wei, Arny A. Ferrando, and Robert R. Wolfe. 2019. "Consumption of a Specially-Formulated Mixture of Essential Amino Acids Promotes Gain in Whole-Body Protein to a Greater Extent than a Complete Meal Replacement in Older Women with Heart Failure" Nutrients 11, no. 6: 1360. https://doi.org/10.3390/nu11061360