Influence of Foliar Kaolin Application and Irrigation on Photosynthetic Activity of Grape Berries


1
Centre for the Research and Technology of Agro-Environmental and Biological Sciences (CITAB), University of Minho, Campus de Gualtar, 4710-057 Braga, Portugal
2
Centre for Environmental and Marine Studies (CESAM), University of Aveiro, Campus de Santiago, 3810-193 Aveiro, Portugal
3
Wageningen Plant Research, Wageningen University and Research Centre (Wageningen-UR), PO Box 16, 6700 Wageningen, The Netherlands
4
Centre of Molecular and Environmental Biology (CBMA), University of Minho, Campus de Gualtar, 4710-057 Braga, Portugal
5
Centre of Biological Engineering (CEB), University of Minho, Campus de Gualtar, 4710-057 Braga, Portugal
*
Author to whom correspondence should be addressed.
Agronomy 2019, 9(11), 685; https://doi.org/10.3390/agronomy9110685
Submission received: 30 September 2019
/
Revised: 23 October 2019
/
Accepted: 24 October 2019
/
Published: 27 October 2019
(This article belongs to the Special Issue Viticulture and Winemaking under Climate Change)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Climate changes may cause severe impacts both on grapevine and berry development. Foliar application of kaolin has been suggested as a mitigation strategy to cope with stress caused by excessive heat/radiation absorbed by leaves and grape berry clusters. However, its effect on the light micro-environment inside the canopy and clusters, as well as on the acclimation status and physiological responses of the grape berries, is unclear. The main objective of this work was to evaluate the effect of foliar kaolin application on the photosynthetic activity of the exocarp and seeds, which are the main photosynthetically active berry tissues. For this purpose, berries from high light (HL) and low light (LL) microclimates in the canopy, from kaolin-treated and non-treated, irrigated and non-irrigated plants, were collected at three developmental stages. Photochemical and non-photochemical efficiencies of both tissues were obtained by a pulse amplitude modulated chlorophyll fluorescence imaging analysis. The maximum quantum efficiency (Fv/Fm) data for green HL-grown berries suggest that kaolin application can protect the berry exocarp from light stress. At the mature stage, exocarps of LL grapes from irrigated plants treated with kaolin presented higher Fv/Fm and relative electron transport rates (rETR200) than those without kaolin. However, for the seeds, a negative interaction between kaolin and irrigation were observed especially in HL grapes. These results highlight the impact of foliar kaolin application on the photosynthetic performance of grape berries growing under different light microclimates and irrigation regimes, throughout the season. This provides insights for a more case-oriented application of this mitigation strategy on grapevines.
1. Introduction
Viticulture is a historically important agronomic and socio-economic sector in Portugal. According to the last report from the International Organization of Vine and Wine (OIV), Portugal is the 11th world and 5th European wine producer [1]. With 14 winemaking regions distributed throughout the country, the Vinhos Verdes or Minho region, as well as the Douro and Alentejo, are the major contributors for national exports and growth of this sector [2].
Grapevine is influenced by a complex and interacting system commonly called terroir, which, according to the OIV [3], includes specific soil, topography, climate, landscape characteristics and biodiversity features, and interaction with applied vitivini-cultural practices. This complex system influences the canopy microclimate and grapevine physiology and development and, consequently, grape berry quality and the organoleptic properties of its wine, which is typical of each region.
Currently, climate change projections point to a particularly pronounced temperature variation, with an overall increase of up to 3.7 °C by the end of this century, compared to the 1985–2005 reference period [4]. These temperature changes will have great impacts in the Mediterranean wine regions [5]. According to recent investigations using very high resolution bioclimatic zoning, both temperature and dryness are predicted to increase in several economically important Portuguese viticulture regions, including the Vinhos Verdes region [6]. Therefore, Portuguese vineyards will be subject to increased stress due to the interaction of the existing high radiation levels with the foreseen elevated air temperature and drought, which, all together, can have high impact on grapevine phenology, physiology, and productivity. Several of these climate impacts have already been reported, such as: earlier phenological timings and shortenings of the grapevine growing season [7], sunburns in leaves and grape berries [8], reduction of stomatal conductance and decrease of photosynthetic rates, either by stomatal and non-stomatal limitations [9], appearance and/or intensification of grapevine-related pests and diseases [10,11], increased grape sugar concentrations that lead to higher wine alcohol levels, lower acidities, and modification of varietal aroma compounds [12], and higher inter-annual yield and wine production variability [13].
In order to mitigate these adverse climate effects, new short-term measures have recently been implemented in Portuguese vineyards, such as smart irrigation [14,15] and foliar application of kaolin [16]. Vineyards are not traditionally irrigated and there are even restrictions on this practice in some regions, such as the Douro region [15]. However, according to a recent projection model, a 10% reduction in grapevine yield is expected in the Minho region if irrigation is not applied [14]. Kaolin is a white, chemically inert, and non-toxic clay material (Al₂Si₂O₅(OH)₄) that can reflect radiation, including photosynthetically active (PAR), ultraviolet (UV), and infrared radiation (IR) [16]. Foliar application of this mineral has become a cost-efficient mitigation strategy to cope with water stress and excessive heat/radiation absorbed by leaves and grape berry clusters, which also proves effective in alleviating negative impacts on grapevines [17,18,19,20,21]. However, the amount and spectral quality of light intercepted by leaves and transmitted/ reflected into the canopy, crucial factors for leaf, and fruit physiology and development [22] are also important aspects to consider when mitigation practices are used.
Previous work done by our group, using pulse amplitude modulated (PAM) chlorophyll fluorescence imaging, has mapped grape berry photosynthesis at a histological level, and revealed both the exocarp and the seed outer integument as the main photosynthetically competent tissues [23]. More recently, we have studied the photosynthetic performance of grape berry tissues from clusters growing at three distinct light microclimates in the canopy and observed microclimate-related differences in their photosynthetic capacity and acclimation status [24]. This led to the hypothesis that, if a specific viticulture practice changes the light reaching the clusters, and alters its light microclimate, it may impact the photosynthetic activity of berry tissues and associated tissue-specific biochemical processes. In fact, foliar kaolin application may have direct implications on light distribution at the whole canopy level, and irrigation is an indirect one, through increased vegetative growth. For instance, it has already been shown that kaolin application generally reduces the photosynthetic rates of individual leaves in other agricultural crops (e.g., apple, almond, and walnut canopies) [25,26], due to a 20–40% increase in the reflection of PAR [27]. However, the photosynthesis of the whole canopy remained unaffected or even increased (9%), because of the better light distribution within the canopy [28,29,30]. In another study, decreased photosynthesis was observed in the inner leaves of irrigated grapevines due to higher vegetative growth [31]. While the function of photosynthesis in fruits is still poorly understood, it can be linked with primary and secondary metabolomic pathways [32,33]. Therefore, any effect on photosynthesis may impact grape berry development and composition.
Therefore, the main objective of the present work was to evaluate the effects of foliar kaolin application on the photosynthetic activity of grape berry tissues from clusters growing at two distinct microclimates, which include high light (HL) and low light (LL) microclimates, of irrigated and non-irrigated grapevines, during the season.
2. Materials and Methods
2.1. Site Description, Applied Treatments, and Sampling
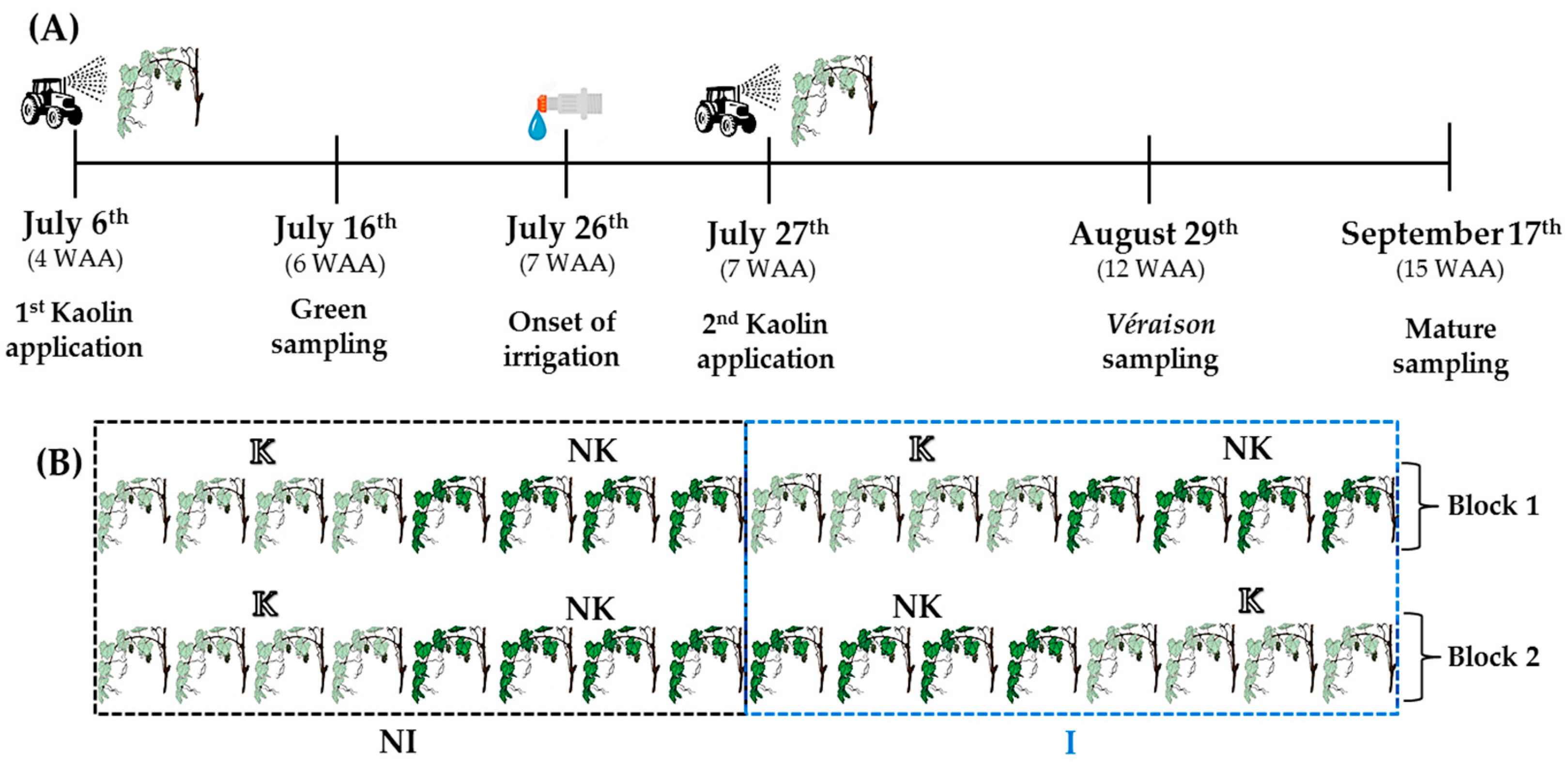
Grape berry samples were collected in 2018 from field-grown ‘Alvarinho’ cultivar grapevines (Vitis vinifera L.) in the commercial vineyard Quinta Cova da Raposa in the Demarcated Region of Vinho Verde, Braga, Portugal (41°34’16.4” N, 8°23’42.0” W). The vineyard is managed by following standard cultural practices applied in organic farming, and is arranged in terraces along a granitic hillside with high drainage. The vine training system applied for this cultivar follows the settings of Sylvoz (Simple Ascending and Recumbent Cord). The sector selected for the trial was located on a hill with NW-SE orientation and the vineyard rows with a NE-SW orientation. The treatments applied were: kaolin (K) and non-kaolin (NK) application on leaves, and irrigation (I) and non-irrigation (NI), in a complete factorial design (four treatment combinations) with two blocks, each with three to four vines per combination treatment (Figure 1B). A suspension of 5% (w/v in water) kaolin (EPAGRO®, Sunprotect, Alverca do Ribatejo, Portugal) was applied twice on leaves on both sides of the rows. On July 6 and 27, corresponding to four weeks after anthesis (WAA) or BBCH-73 (BBCH-scale used for grapes by Lorenz et al. [34]) and seven WAA or BBCH-77, respectively. Irrigation of half of the plants, started on July 26 (seven WAA, BBCH-77), (Figure 1A,B). Water was applied by drip irrigation with one dripper per vine and a drip line placed approximately 80 cm above the soil. Irrigation occurred every three days, once a day either early in the morning or late in the afternoon, for 2 h, with an average dripper capacity of 5.5 ± 1.6 L h−1 (n = 12 randomly selected drippers, ± SD). Clusters with contrasting light exposure were also selected to harvest grape berries during their development. These were called low light (LL) and high light (HL) clusters. LL clusters grew in the shaded inner zones of the canopy, which were exposed only to diffuse, reflected, and transmitted light, while HL clusters were exposed to direct or reflected sunlight most of the day. Six independent sub-clusters (three per block), each containing 15–20 grape berries, were collected randomly from clusters growing at each of the experimental conditions (four treatments × 2 microclimates) from the southeast side of rows. Berries were harvested in the morning (9–10 a.m.) at three distinct developmental stages: Green (16 July, 6 WAA, BBCH-75), Véraison (29 August, 12 WAA, BBCH-83), and Mature (17 September, 15 WAA, BBCH-89). The material was transported in refrigerated boxes to the Center for Environmental and Marine Studies (CESAM) laboratory and used within 2–6 h for imaging fluorometry experiments. For other assays, berries were immediately frozen in liquid nitrogen and stored at −80 °C.
2.2. Light Intensity and Temperature Measurements for Microclimate Characterization
In order to characterize the microclimates in the vicinity of the growing clusters of all experimental conditions, light intensities and temperatures were registered on cloudless days (mean of 1500 ± 300 µmol photons m−2 s−1), between 15 h and 17 h, at green and mature stages, as described by Garrido et al. [24]. The light intensity (µmol photons m−2 s−1) was measured with a radiometer (LI-COR, LI-250 Light Meter, Lincoln, NE, USA) and the temperature (°C) was measured with an infrared digital thermometer (Infrared, DT8380, Beijing, China). Both parameters were determined in the frontal region of the clusters (LL and HL), at the southeast side of the row, and in full sun-exposed leaves. The devices were placed perpendicularly to the plant organ (cluster or leaf). The light sensor was placed on the organ surface facing outward, which registered the light intensity reaching at this point, and the thermometer was pointing to the organ at a distance of about 15 cm, which registered an average organ temperature. Sixteen replicate measurements per treatment were considered on randomly selected vines.
2.3. Kaolin Film Transmittance and Reflectance
Transmittance and reflectance spectra were obtained using a spectrometer (USB2000-VIS-NIR, grating #3, Ocean Optics, Duiven, The Netherlands), connected to a 400 mm-diameter fiber optic (QP400-2-VIS/NIR-BX; Ocean Optics), and recorded using the spectral acquisition software Spectra Suite (Ocean Optics, https://oceanoptics.com/). The transmittance spectrum was obtained by spreading a 5% (w/v) kaolin suspension prepared in 70% ethanol (allowing fast evaporation to prevent solvent interference), over a glass plate, which simulated particle distributions similar to those observed in the field. A halogen lamp was placed underneath to illuminate the spectrometer sensor positioned 3 cm above the glass plate. Different spectra were obtained from different areas randomly (n = 3). The reflectance spectrum was obtained, according to Dinis et al. [18], by pointing the fiber optics perpendicularly to the surface of collected leaves illuminated by the same halogen lamp. Three independent spectra were obtained from different leaf regions of both kaolin-treated and non-treated leaves. Transmittance and reflectance spectra were recorded for the 370–900 nm spectral range, with a spectral resolution of 0.33 nm. The transmittance spectra were expressed as a percentage of the controls (glass). Reflectance spectra were normalized to the spectrum reflected from a reference white panel (WS-1-SL Spectralon Reference Standard, Ocean Optics).
2.4. Chlorophyll Fluorescence Analysis
The photosynthetic activity of grape berry tissues was assessed as described by Garrido et al. [24]. For this, an imaging chlorophyll fluorometer was used (Open FluorCAM 800 MF; Photon Systems Instruments, Drásov, Czech Republic), which was comprised of four 13 × 13 cm LED panels emitting red light (emission peak at 621 nm, 40-nm bandwidth) and a 2/3 inch CCD camera (CCD381, Beijing, China) with a F1.2 (2.8–6 mm) objective. Two of the LED panels provided modulated measuring light (<0.1 μmol m−2 s−1) and the other two provided saturating pulses (>7500 μmol m−2 s−1, 0.8 s). Chlorophyll fluorescence images were captured and processed using FluorCam7 software (Photon Systems Instruments, Drásov, Czech Republic).
In a dark cabinet, exocarps and seeds were separated from dark-adapted berries and disposed in 8 × 8-well plates filled with water. Two independent plates were prepared for each microclimate (LL and HL), with each comprising all treatments and tissues. Exocarps and seeds were placed alternately in three rows each, using two columns per treatment, in a total of 12 biological replicates per condition and tissue (n = 3 × 2 × 2 = 12). Each plate was subjected to the experiments described below.
The maximum quantum efficiency of photosystem II [Fv/Fm = (Fm − F0)/Fm], which is a chlorophyll fluorescence parameter that reflects the probability of electrons being transferred from the PSII reaction center for the transport chain of electrons by quanta absorbed [35,36], was computed following a saturation pulse (SP). The isolated tissues were then acclimated to an actinic light (AL) of 200 µmol photons m−2 s−1 for 15 min, and, after a new SP, the effective quantum yield of PSII [ΦII = (F′m − Fs)/F′m] was computed. This parameter correlated with the quantum yield of CO2 fixation in a wide range of physiological conditions [37]. From ΦII and PFR (photosynthetic photon fluence rates) (200 µmol photons m−2 s−1), the relative electron transport rate through PSII (rETR200 = ΦII × PFR) was calculated. Then, tissues were exposed to 1500 µmol photons m−2 s−1 for 15 min, with an SP being applied every 3 min. The last F’m values (at 15 min) were used to calculate the non-photochemical quenching [NPQ = (Fm − F’m)/F’m].
2.5. Analysis of Chlorophylls and Carotenoids by High Performance Liquid Chromatography Coupled to A Photodiode Array Detector (HPLC-PDA)
The extraction procedure was adapted from Fraser et al. [38]. Freeze-dried material (20 mg) of grape berry tissues, which includes exocarp and seed, was extracted in 1.8 mL of chloroform/methanol (1:1) (chloroform - Emsure®, Darmstadt, Germany, methanol - Biosolve®, Dieuze, France) with both 0.1% (w/v) butylated hydroxytoluene (BHT, Sigma®, Zwijndrecht, The Netherlands) as an antioxidant and Sudan 1 (0.5 μg mL−1) as the internal standard (IS). The samples were vortexed (10 s), kept on ice for 30 min (vortexed in between), and then sonicated for 15 min (Branson®, 3510 Ultrasonic Cleaner, Danbury, CT, USA). These steps were performed twice. After that, the samples were centrifuged at 16,100× g (Eppendorf®, Centrifuge 5415 R, Hamburg, Germany) and the supernatant (approx. 1200 μL) was transferred to a new Eppendorf tube with a perforated lid. The samples were dried for 1 h in a Speedvac (Savant®, SC100, Schiphol, The Netherlands) and then stored at −80 °C until the next steps. Prior to high performance liquid chromatography (HPLC) analysis, samples were dissolved in 200 μL ethylacetate solution containing 0.1% (w/v) BHT, sonicated (10 min), and then centrifuged as above. Samples were protected from light and kept on ice during all of these procedures. The supernatant (180 μL) was transferred to amber-colored 2 mL HPLC vials with a glass insert and sealed.
The HPLC-PDA procedure was adapted from Mokochinski et al. [39]. The samples (20 μL) were analyzed using an HPLC (Waters Alliance e2695 Separations Module, Milford, MA, USA) coupled to a photodiode array detector (PDA) (Waters 2996) over the 240 to 700 nm UV/Vis range. Separation was performed on a reverse-phase C30 column (250 × 4.6 mm i.d., S-5 μm - YMC Carotenoid, Komatsu, Japan) kept at 35 °C with a flow rate of 1.0 mL min−1. The compounds were identified based on comparisons of retention times and absorption spectra (240 to 700 nm) with authentic standards.
2.6. Statistical Analysis
Results were statistically analyzed using Analysis of Variance tests (two-way ANOVA), followed by post hoc multiple comparisons using the Bonferroni test whenever the factors had significant effects (GraphPad Prism version 5.00 for Windows, GraphPad Software, La Jolla, CA, USA). Significant differences (p ≤ 0.05) between sample groups are indicated with different letters. Notation with an asterisk means that only one factor (kaolin or irrigation) was significant.
3. Results and Discussion
3.1. Climatic Conditions and Microclimate Characteristics
In order to characterize the climate during the growing season at the study site (Braga), we used the official information available from the Instituto Português do Mar e da Atmosfera (IPMA) [40], to determine the temperatures and total precipitation during 2018 (Figure S1). This growing season was atypical from a climatic point of view, with a relatively cold and extremely dry winter, which caused a delay of sprouting/flowering for two to three weeks [41], and a relatively cold spring with rainy periods during the vegetative growth of the grapevines.
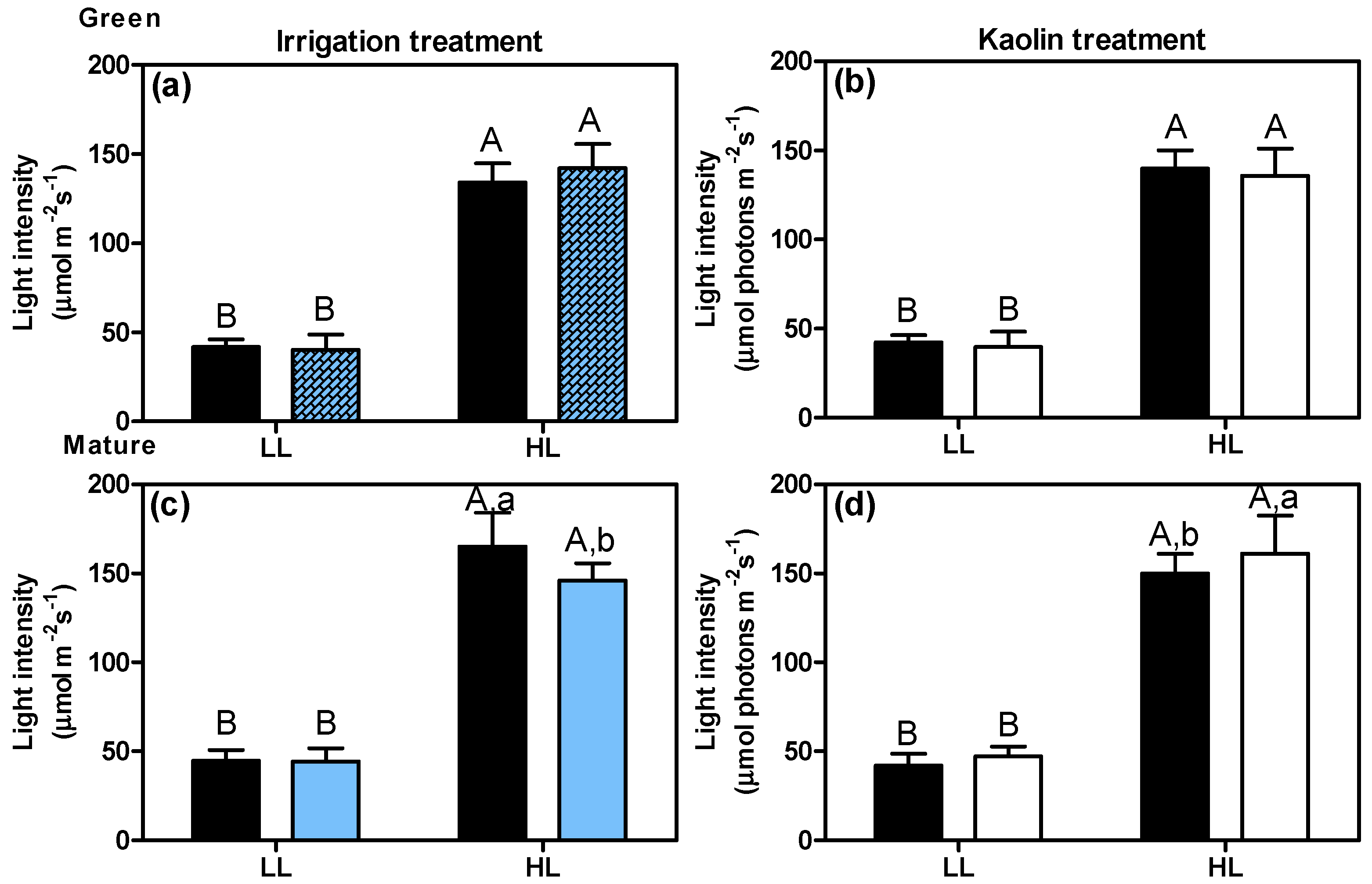
To characterize the microclimates for the LL and HL berry clusters (two a priori selected distinct light microclimates within the canopy), measurements of light intensities and temperatures were performed at the cluster level, at two time points during the growing season, i.e., when the berries were still green (green stage) and, two months later, when the berries were at their mature stage of ripening (Figure 2 and Figure 3). Figure 2 depicts the average light intensities at LL and HL clusters growing under the different experimental conditions: i.e., irrigation/non irrigation (Figure 2a,c) and with kaolin/without kaolin (Figure 2b,d). The two microclimates were clearly distinct at both ripening stages, with HL clusters receiving about three-fold more light than LL clusters. At the green stage, i.e., before the onset of irrigation (Figure 1A), no significant differences were detected between the two sets of plots assigned to the subsequent irrigation experiment (four plots for irrigated (I) plants, i.e., 2 × NK-I and 2 × K-I) and four plots for non-irrigated (NI) control plants (2 × NK-NI and 2 × K-NI), see Figure 1B) (Figure 2a), which reveals that there were no plot-related effects on microclimate light intensity. At this early ripening stage, and with the adopted measurement procedure, no differences were detected with respect to light intensities reaching the berry clusters due to foliar kaolin application (Figure 2b). However, at a mature stage, both irrigation and kaolin had a small but significant effect on the light intensity reaching the HL clusters (Figure 2c,d). Irrigation slightly reduced the light intensity, likely due to the better vegetative growth of the plants, while foliar kaolin application increased it, likely due to an increased light reflection to both the interior and lower levels of the canopy. In the LL clusters, these effects of irrigation and kaolin on light intensity were not observed, at this time of day.
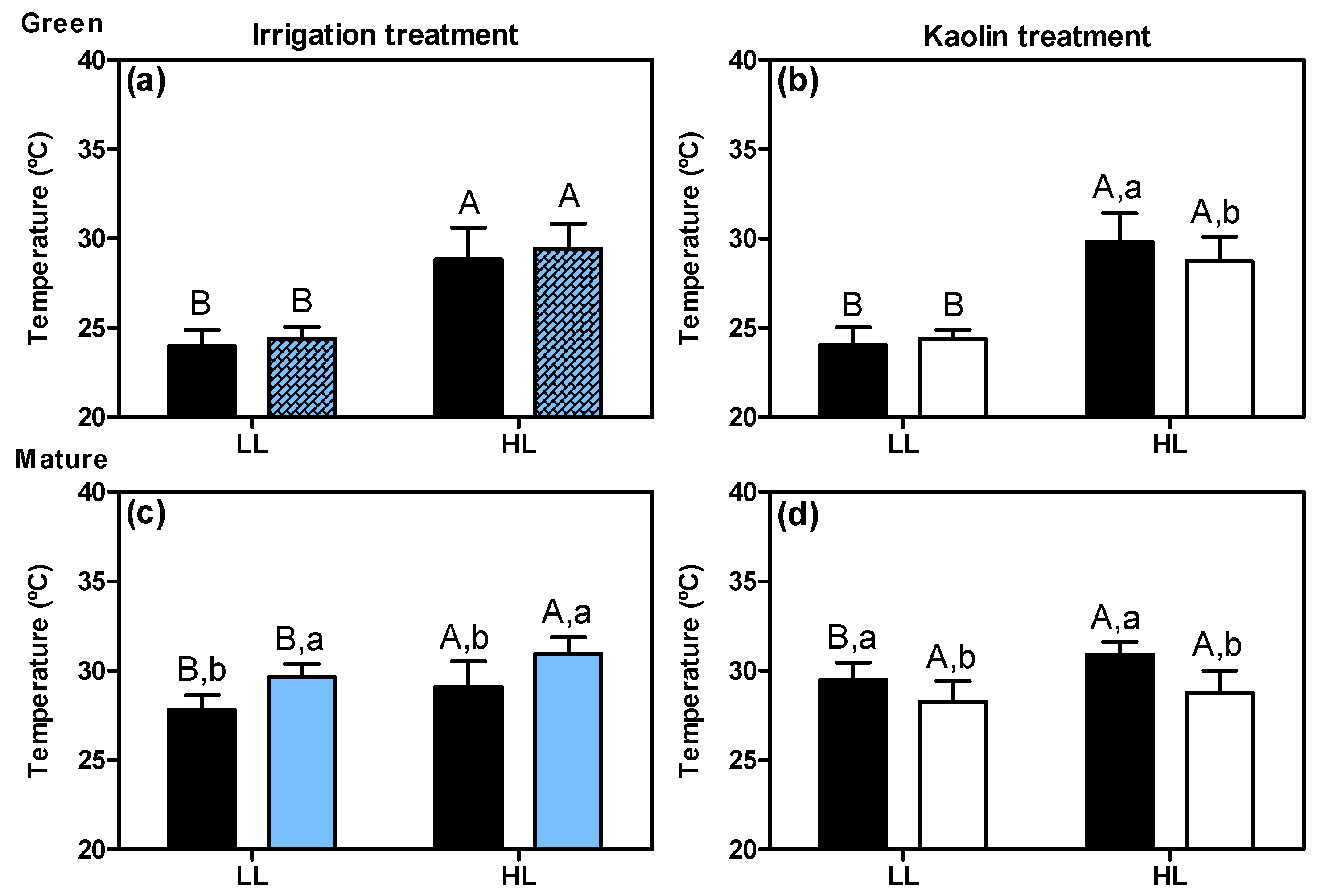
HL grapes consistently experienced higher temperatures than LL ones (Figure 3), and both I and K treatments exerted significant and contrasting effects on this microclimate parameter, mainly at the mature stage. Again, and consistent with what was observed and discussed above for light intensity, no effect was detected for I treatment on the grape berry temperature at the green stage (before the onset of irrigation) (Figure 3a).
The Kaolin application led to a significant decrease in the temperature of the HL clusters at both green and mature stages (e.g., at this latter stage—from 31 °C to 28.7 °C), and of the LL clusters at the mature stage only (e.g., from 29.5 °C to 28 °C) (Figure 3b,d). The fact that LL clusters’ temperature at the green stage were not affected by kaolin was likely related to the relatively low air temperature at this time of the growing season (early July, Figure S1). Thus, kaolin application on the leaves may increase the incident light by increasing the light reflection inside the canopy (Figure 2d), while maintaining a cooler microclimate for the growing berries, especially during the hot summer days, independently of the irrigation regime. This demonstrates one of the advantages of this mitigation strategy at the grape berry level. The kaolin solution sprayed on the leaves also resulted in leaf temperature reduction (Figure S2d). The present results are in agreement with previous studies in both grapevine leaves and berries [42,43], as well as other crops [26,27]. Therefore, it is likely that kaolin applied to leaves provides cooler temperatures throughout the grapevine by reducing the total amount of radiation transmitted into the canopy. This is also shown by thermal imaging in apple trees [30]. Furthermore, different training systems of the vineyard might influence the light intensities and temperatures inside the canopy [44,45]. For instance, the vine canopy was denser in our previous study in 2015 [24], which resulted in an LL microclimate characterized by much lower light reaching the clusters compared to that in the present study, with major impacts on grape berry photosynthetic competence.
At the mature stage, plant irrigation resulted in a significant increase in the grape temperature of both LL and HL clusters (Figure 3c). This increase was unexpected, since a previous study reported lower berry temperature as a result of irrigation, rather than a higher temperature [46]. This response is very interesting and clearly, additional studies, which are controlling/measuring the soil temperature in the rhizosphere, are required to determine the effect of irrigation and the irrigation procedure on the temperature of grape clusters.
3.2. Kaolin Film Transmittance and Reflectance Spectral Properties
To characterize the potential effects of kaolin applied to the leaves with regard to light intensity and quality, we performed transmittance and reflectance studies. For this purpose, the transmittance spectrum of a film of kaolin solution (5% w/v) on a glass plate was determined (Figure 4a). Although a high percentage of most photosynthetically active radiation (PAR) wavelengths is transmitted by the kaolin film, the blue light range is the least transmitted. The reflectance spectra obtained for leaves of grape plants sprayed with and without kaolin are represented in Figure 4b. Our results showed a relevant percentage of PAR is reflected by this white mineral, as compared to non-spayed (NK) vine leaves, rather than exclusively or mainly reflecting in the ultraviolet (UV) and infrared radiation (IR) ranges [47]. Additionally, kaolin was more efficient in reflecting UV light than IR light (in the measured ranges). These results are in accordance with previous studies using grapevine leaves [18] and other crops [27,28,48,49]. Thus, the beneficial effect of kaolin application is related to the reflection of excess radiation outwards, which reduces the risk of light stress-induced damage to leaves and fruit [47], while transmitting a very significant proportion of PAR.
Together, these results call attention to the fact that foliar kaolin application may directly impact the photosynthesis of the sprayed leaves but also have an indirect effect on the non-sprayed leaves and grape berry clusters inside the canopy.
3.3. Effects on Berry Photosynthesis and Photosynthetic Pigments
3.3.1. Maximum Quantum Efficiency of PSII
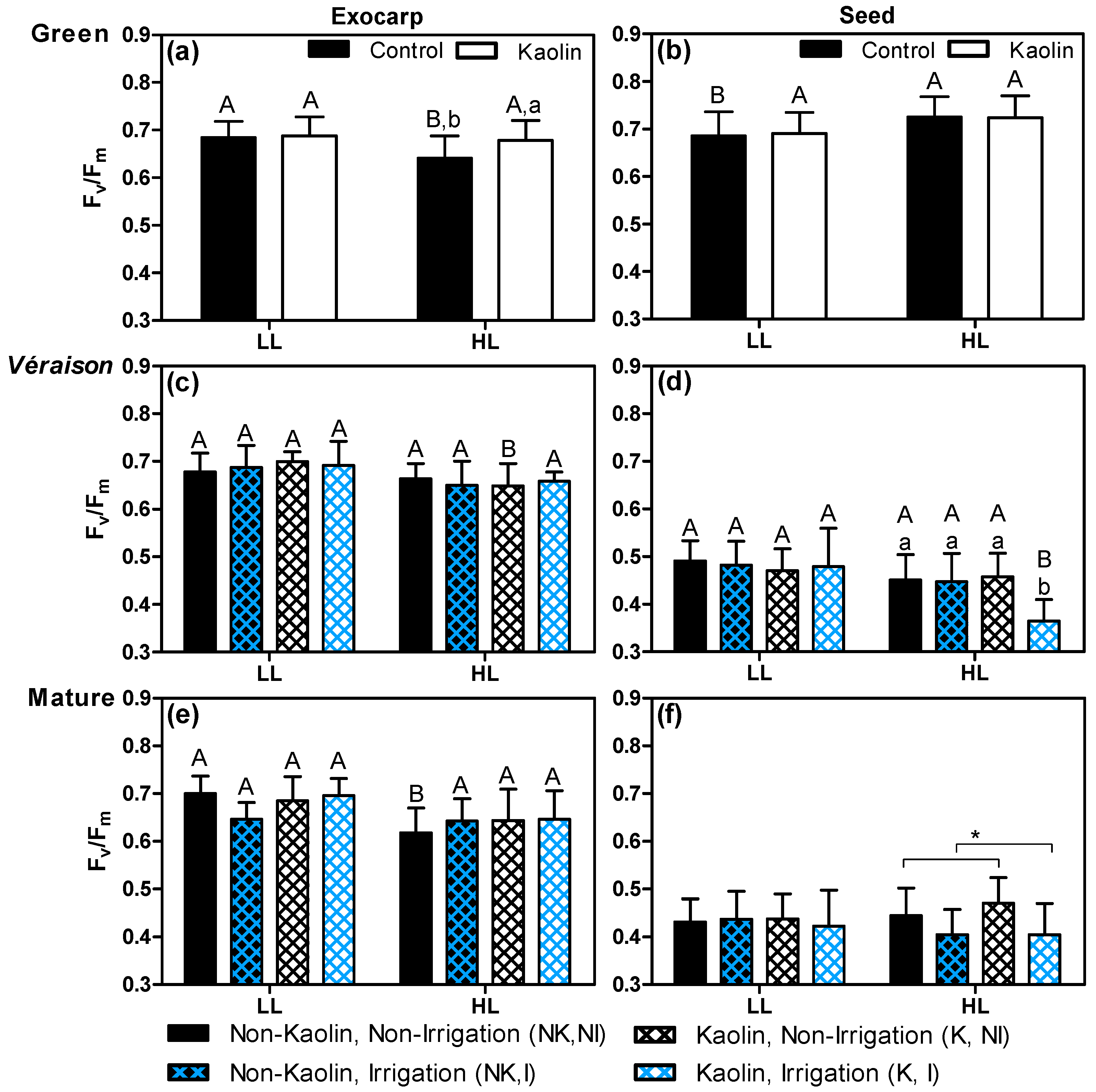
The maximum quantum efficiency of PSII (Fv/Fm) was determined ex planta under controlled conditions, using both exocarps and seeds from grape berries grown under the different treatments, microclimates, and three ripening stages (Figure 5). At the green stage, Fv/Fm was similar in both berry tissues (Figure 5a,b). Upon further ripening, the exocarp kept its Fv/Fm values (~0.7), while the seeds showed a significant decrease in this parameter, which reached values around 0.4–0.5 at the mature stage, which was in accordance with what was reported by Garrido et al. [24].
The cluster microclimate (LL vs. HL) had a significant effect on the Fv/Fm values in both tissues and all developmental stages, with the exception of seeds at the mature stage, but exocarps and seed integuments responded differently to a light microclimate [24]. At the green stage, the exocarps from berries under control conditions, showed lower Fv/Fm values in HL clusters, while their seeds showed significantly higher values than those from LL berries (Figure 5a,b). This was likely due to their inner location where the light transmitted through the skin and flesh tissues reaches values as low as 2% of the incident photon flux density (PFD) [50], which eventually translates a light limitation effect in LL clusters. These microclimate effects were more or less maintained in exocarps upon subsequent ripening (see NK + NI Figure 5a,c,e, while not being significant in véraison), while, in seeds, the difference between LL and HL clusters disappeared (Figure 5b,d,f), likely related to the (large) intrinsic ripening-dependent decrease in photosynthetic competence of this tissue.
Both kaolin and irrigation treatments of plants differentially influenced the Fv/Fm values of the two berry tissues, with the seeds globally more responsive than exocarps, particularly to the irrigation treatment, which induced a significant effect at véraison and mature stages (Figure 5d,f). On the other hand, at these latter stages, no effects from treatments were detected on the Fv/Fm of exocarps. This tissue only responded to kaolin treatment at the green stage, where HL berries showed an increment (6%) on Fv/Fm by kaolin application (Figure 5a). In fact, the decrease in Fv/Fm values of exocarps when comparing LL with HL berries at a green stage in control conditions (Figure 5a) had already been observed [24], which revealed that microclimates with higher luminosity can decrease Fv/Fm values of exocarps at this stage. Together, these results suggest that foliar kaolin may protect the berry exocarp from excess light at a stage when the grape berry photosynthetic phenotype is still developing [24]. In HL seeds, the most prominent effect was a decrease in Fv/Fm in irrigated-treated plants (Figure 5d,f). However, at véraison, this effect was observed only in kaolin-treated plants (Figure 5d). This apparent paradoxical effect may be related with the increased temperatures observed in irrigated grape berries in the hottest months (Figure 3c and Figure S1). In fact, the inhibitory effect of higher temperatures on Fv/Fm was already reported for grapevine leaves [51].
3.3.2. Relative Rate of Electron Transport Through PSII (rETR200)
The relative electron transport rate through photosystem II (rETR) was determined after acclimation of the berry tissues to an AL intensity of 200 µmol photons m−2 s−1 (rETR200) in order to simulate average field light conditions.
At the green stage, the rETR200 was higher in HL grape berries than in LL berries (both in control and kaolin-treated plants), especially in seeds (Figure 6a,b). Interestingly, a positive influence of kaolin application was observed in LL exocarps at this stage (Figure 6a), which suggests that more PAR reflected by kaolin is reaching the inside of the canopy, which may improve the exocarp photosynthesis of berries in shaded microclimates. At the véraison stage, this positive effect of kaolin was not detected, while it was evident at the mature stage (Figure 6e). In addition, a clear decrease in rETR200 was observed in seeds, especially in HL clusters (p < 0.0001) during ripening (Figure 6b,d,f), which is similar to the results obtained for Fv/Fm. In addition, in seeds from HL-grown berries at the véraison stage of irrigated plants, the kaolin treatment led to a significant decrease in rETR200 (Figure 6d: NK, I compared to K, I), which was observed for the parameter Fv/Fm (Figure 5d). This pointed to a possible effect of berry temperature on seed photosynthesis. No reports were found for the effect of temperature on grape berry photosynthesis, but a continuous four-day exposure to high temperatures (38–40 °C) led to a decrease in photosynthetic activity of grapevine leaves [52,53,54].
3.3.3. Non-Photochemical Quenching
A major component of non-photochemical quenching (NPQ) is the primary protective mechanism against light-induced photoinhibition, which involves various processes dissipating excessive non-radiative energy [55], including the xanthophylls cycle (e.g., as shown in grapevines, [56]) and phosphorylation/dephosphorylation of light harvesting complexes [57].
The NPQ results are represented in Figure 7. When comparing the berry tissues, it can be concluded that the exocarp tissue consistently exhibits roughly two-fold higher NPQ values than seeds. This result suggests that the exocarp exhibits more developed mechanisms of photoprotection, which is consistent with the fact that it is an external, more exposed tissue. During berry ripening, NPQ values decreased in both tissues, especially in seeds. Seeds attained very low NPQ values at later stages, which is in line with their Fv/Fm (Figure 5b,d,f) and rETR200 (Figure 6b,d,f) profiles, which likely reflect the normal ripening-related loss of photosynthetic functioning of seeds. For the exocarps, dissipation or quenching mechanisms other than NPQ may explain the result, since this tissue maintains high photosynthetic activity until the mature stage (Figure 6a,c,e). Accumulation of carotenoids in white berries (‘Sauvignon Blanc’) was also increased in response to increasing levels of solar light in the canopy, which shows that the berries utilize these photosynthesis-related pigments in photo-acclimation responses and/or as “sunscreens” [58].
Regarding the effect of treatments, we found that the foliar kaolin application promoted lower NPQ in HL exocarps at the green stage, when compared with their NK controls (Figure 7a), which suggests that kaolin helps to protect these HL berries from excessive radiation absorption. This is similar to the Fv/Fm results (Figure 5a).
At the véraison stage, both HL exocarps and HL seeds showed increased NPQ in irrigated-treated grape berries (Figure 7c,d). In LL exocarps, NPQ was also increased in K,I berries, when compared to the remaining treatment combinations (Figure 7c). This increase in NPQ values of grape berries in irrigated plants, was lost at the mature stage, even though it is important to note that NPQ values were already very low at this stage (Figure 7e,f). This irrigation-related feature had already been observed with other parameters and is discussed above. The increased temperatures registered at later developmental stages of grape berries (Figure 3c), can impose other limitations or impairments, and, thus, recruit more energy-dissipation by NPQ, and eventually by other dissipative mechanisms.
3.3.4. Photosynthetic Pigments
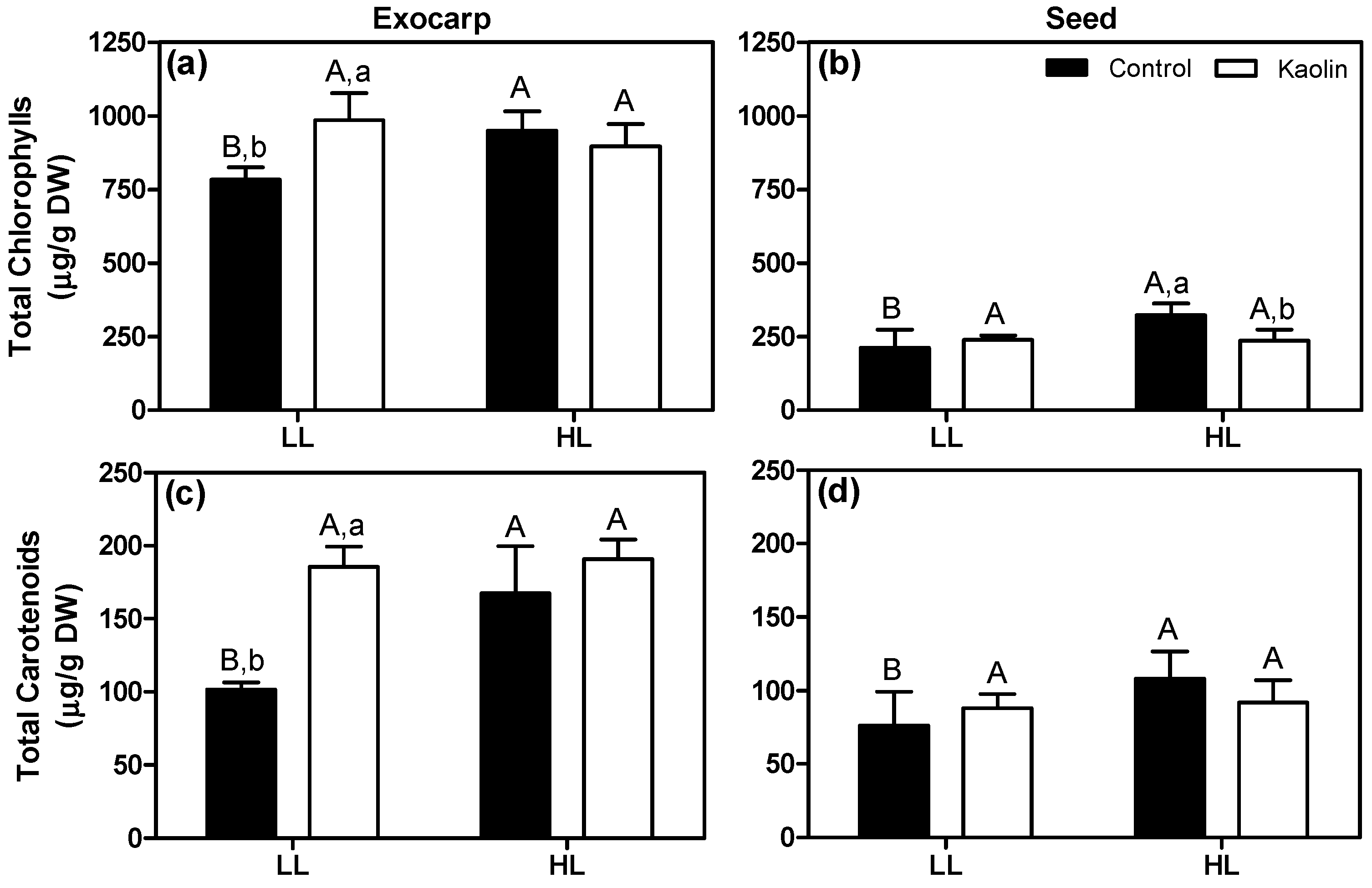
To better evaluate the impact of foliar kaolin application on the light microclimate of grape berry clusters and its relationship with berry photosynthesis, and non-photochemical mechanisms, photosynthetic pigments were quantified in exocarps and seeds of both LL and HL-exposed grapes. Results obtained for the green stage are depicted in Figure 8 (for later stages, see Supplementary Materials). At control conditions, the HL berries had higher levels of both chlorophylls and carotenoids than LL berries, in both tissues. Additionally, and in line with the rETR200 results (Figure 6a), kaolin application resulted in a marked increase by 26% in chlorophylls and 82% in carotenoids content in exocarps from LL berries (Figure 8a,c), which support the idea that more light reached the inner parts of the kaolin-sprayed canopy. This is fundamental to build the photosynthetic machinery [59]. During ripening, the photosynthetic pigments decrease in both tissues and especially in the seed integuments (Figures S3 and S4) and no consistent and conspicuous effects by combined mitigation treatments were observed (Figure S3).
In addition, and supporting the view discussed above, the higher grape berry temperature was registered in irrigated treatments at later developmental stages (Figure 3c), by imposing physiological impairments. The temperature recruits more energy-dissipation by NPQ (Figure 7), which is the fact that carotenoids contents (Figure S4), but not chlorophylls (Figure S3), were also increased by irrigation treatment, for both tissues at the véraison stage.
Overall, the results obtained by pulse amplitude modulated fluorometry showed that, for the external tissue, exocarp, and foliar kaolin application led to an increase of Fv/Fm (Figure 5a, HL), rETR200 (Figure 6a,e LL), and a reduction in non-photochemical quenching (Figure 7a, HL). To our best knowledge, this is the first work assessing the impact of foliar kaolin application on photochemical and non-photochemical functions in grape berries. Recently, it was verified that grapevine leaves with kaolin display the same response, i.e., an increase in Fv/Fm, Φll, and ETR, and a decrease in NPQ [18,60]. Similar results were also reported for olive leaves [61]. In this way, and in terms of photochemical processes, those kaolin-treated leaves have lower photo-inhibitory damage [17,62], and the open PSII reaction centers captured the light absorbed by PSII antenna more efficiently [17,35]. This response was likely due to a reduced loss of excitation energy by thermal dissipation, which could compete with its transference to PSII reaction centers, as shown by the lower NPQ values [17,35].
For exocarps of grape berries growing in inner parts of the canopy (LL microclimate), the photosynthetic results revealed that foliar kaolin application, may cause an extra “sunscreen” effect, and did not have a negative effect on those parameters, which we conjectured in our previous work [24]. The increased reflection provided by this mineral to inner parts of the canopy allowed good photochemical performance of LL exocarps, which is contrary to what we hypothesized in our previous work [24]. This contributes to higher carbon gains at the whole canopy level and also at the fruit level.
Regarding the results for the seed integument (internal organ), the positive effects of kaolin were observed mainly in non-irrigated plants such as an increase in Fv/Fm (Figure 5f, HL) and a decrease in NPQ (Figure 7d, HL). In more temperate or Mediterranean regions, this seems like a positive effect, but these results also show the importance of the irrigation system. The interaction between kaolin application and irrigation treatments on grapevine leaves have been studied before [42,63,64,65,66]. However, based on our knowledge, no study has approached the impacts on photosynthetic activity at the grape berry level, using chlorophyll fluorescence analysis.
4. Conclusions
The purpose of the current study was to assess the effects of foliar application of kaolin and irrigation, as abiotic stress mitigation strategies, on the photosynthetic activity of exocarps (skins) and seeds of grape berries growing under different light microclimates in the canopy. One of the most relevant findings was that the kaolin applied to leaves increased the photosynthetic activity of both exocarps and seed integuments of berries growing under low light conditions in the canopy. This is likely due to higher reflection of PAR to the inner zones. We believe, though, that the beneficial effects will depend on the canopy structure and on the incident radiation, with denser canopies and higher radiations conferring higher overall photosynthetic gains. Somewhat puzzling was the observation that seeds of irrigated plants showed lower photosynthetic activities, in the véraison and mature stages, especially under kaolin treatment. Several causes may explain this unexpected phenomenon, so more detailed and ad-hoc design studies should be conducted to address this relevant finding.
This comprehensive study provides the first evidence of foliar kaolin application as a procedure allowing the modulation of photosynthesis in the grape berry, but also calls attention to the importance of the irrigation system. In this way, this knowledge can be used by farmers to support their decisions concerning sustainable adaptation strategies applied on vineyards. Research to unveil the function of berry tissues’ photosynthesis on the metabolome of the grapes is already underway, which ultimately contributes to the final quality of the fruit and wine.
Supplementary Materials
The following are available online at https://www.mdpi.com/2073-4395/9/11/685/s1. Figure S1. Meteorological elements from IPMA Institute from Braga city. (a) Temperature (°C) maximal, average, and minimal. (b) Total precipitation (mm). Figure S2. Temperatures of full exposed leaves at the green stage (a,b) and the mature stage (c,d), for plants with irrigation (blue columns, note: the textured blue columns at green stage i.e., before the onset of irrigation, represent the measurements in the plots that were later irrigated) and foliar kaolin application (white columns). Black columns correspond to the respective controls. Values represent means with standard deviation (n = 16 plants). Statistical notation: per ripening stage, different lowercase letters refer to significant differences (p ≤ 0.05) between treatments. Whenever letters are omitted, it means that the respective factor did not have a significant effect. Figure S3. Chlorophylls concentration mean values (n = 3, +SD) of exocarp and seed obtained from LL and HL grape berries grown under the four combinations of the two treatments applied: irrigation (I)/ non-irrigation (NI) × kaolin (K)/ non-kaolin (NK). Samples were collected at three development stages (green, véraison, and mature). Statistical notation: for each developmental stage, capital letters refer to differences between light microclimates within the same treatment combination, and lowercase letters refer to differences between treatment combinations within each light microclimate (mean values with a common letter were not significantly different). When capital and lowercase letters are omitted, the respective factor did not have a significant effect (two-way ANOVA p > 0.05). Figure S4. Carotenoids concentration mean values (n = 3, +SD) of exocarp and seeds. All the microclimate conditions, treatment combinations, and statistical information are the same as in Figure S3.
Author Contributions
Conceptualization, A.G., A.C. (Artur Conde), R.V., and A.C. (Ana Cunha) Methodology, A.G., J.S., and A.C. (Ana Cunha) Formal analysis, A.G. and A.C. (Ana Cunha). Investigation, A.G. and A.C. (Ana Cunha). Resources, J.S. and R.V. Writing—original draft preparation, A.G. Writing—review and editing, A.C. (Ana Cunha), J.S., R.V., and A.C. (Artur Conde). Supervision, A.C. (Artur Conde), R.V., and A.C. (Ana Cunha) Project administration, A.C. (Ana Cunha).
Funding
The FCT-Portuguese Foundation for Science and Technology by the grant provided to Andreia Garrido (PD/BD/128275/2017), under the Doctoral Program “Agricultural Production Chains – from fork to farm” (PD/00122/2012), funded this research and APC.
Acknowledgments
The National Funds by FCT - Portuguese Foundation for Science and Technology, under the strategic programmes UID/AGR/04033/2019 and UID/BIA/04050/2019, and the project “INTERACT - VitalityWine - NORTE-01-0145-FEDER-000017 – funded by Norte2020 supported the work. The FCT and FEDER/COMPETE/POCI - Operational Competitiveness and Internationalization Program, under Project the projects MitiVineDrought – PTDC/BIA-FBT/30341/2017 (POCI-01-0145-FEDER-030341), and POCI-01-0145-FEDER-006958 also supported this work. Artur Conde was supported with a post-doctoral fellow of the mentioned INTERACT/VitalityWine project with the Reference BPD/UTAD/INTERACT/VW/218/2016, and also supported by a post-doctoral researcher contract/position within the project “MitiVineDrought” (PTDC/BIA-FBT/30341/2017 and POCI-01-0145-FEDER-030341). This work also benefited from the networking activities within the European Union-funded COST Action CA17111 – “INTEGRAPE - Data Integration to maximize the power of omics for grapevine improvement”. Authors acknowledge the owner from Quinta Cova da Raposa, Manuel Taxa, who provided the samples, Susana Chaves (from CBMA) for her English grammar revision, and also all support given by the Biology Department of the School of Sciences from the University of Minho.
Conflicts of Interest
The authors declare no conflict of interest.
References
- International Organisation of Vine and Wine Statistical Report on World Vitiviniculture; OIV: Paris, France, 2018.
- Lavrador da Silva, A.; João Fernão-Pires, M.; Bianchi-de-Aguiar, F. Portuguese vines and wines: Heritage, quality symbol, tourism asset. Ciência Técnica Vitivinícola 2018, 33, 31–46. [Google Scholar] [CrossRef]
- OIV. Definition of Vitivinicultural “Terroir”; OIV: Paris, France, 2010. [Google Scholar]
- IPCC (International Panel on Climate Change). Special Report on Climate Change, Desertification, Land Degradation, Sustainable Land Management, Food Security, and Greenhouse gas fluxes in Terrestrial Ecosystems. Summary for Policymakers; IPCC: London, UK, 2019. [Google Scholar]
- Ferrise, R.; Moriondo, M.; Trombi, G.; Miglietta, F.; Bindi, M. Climate Change Impacts on Typical Mediterranean Crops and Evaluation of Adaptation Strategies to Cope With. In Advances in Global Change Research; Springer: Dordrecht, The Netherlands, 2013. [Google Scholar]
- Fraga, H.; Malheiro, A.C.; Moutinho-Pereira, J.; Jones, G.V.; Alves, F.; Pinto, J.G.; Santos, J.A. Very high resolution bioclimatic zoning of Portuguese wine regions: Present and future scenarios. Reg. Environ. Chang. 2014, 14, 295–306. [Google Scholar] [CrossRef]
- Fraga, H.; Santos, J.A.; Moutinho-Pereira, J.; Carlos, C.; Silvestre, J.; Eiras-Dias, J.; Mota, T.; Malheiro, A.C. Statistical modelling of grapevine phenology in Portuguese wine regions: Observed trends and climate change projections. J. Agric. Sci. 2016, 154, 795–811. [Google Scholar] [CrossRef]
- Van Leeuwen, C.; Darriet, P. The Impact of Climate Change on Viticulture and Wine Quality. J. Wine Econ. 2016, 11, 150–167. [Google Scholar] [CrossRef] [Green Version]
- Moutinho-Pereira, J.; Magalhães, N.; Gonçalves, B.; Bacelar, E.; Brito, M.; Correia, C. Gas exchange and water relations of three Vitis vinifera L. cultivars growing under Mediterranean climate. Photosynthetica 2007, 45, 202–207. [Google Scholar] [CrossRef]
- Caffarra, A.; Rinaldi, M.; Eccel, E.; Rossi, V.; Pertot, I. Modelling the impact of climate change on the interaction between grapevine and its pests and pathogens: European grapevine moth and powdery mildew. Agric. Ecosyst. Environ. 2012, 148, 89–101. [Google Scholar] [CrossRef]
- Bois, B.; Zito, S.; Calonnec, A.; Ollat, N. Climate vs grapevine pests and diseases worldwide: The first results of a global survey. J. Int. Des Sci. La Vigne Du Vin 2017, 51, 133–139. [Google Scholar] [CrossRef]
- Mira de Orduña, R. Climate change associated effects on grape and wine quality and production. Food Res. Int. 2010, 43, 1844–1855. [Google Scholar] [CrossRef]
- Cunha, M.; Richter, C. The impact of climate change on the winegrape vineyards of the Portuguese Douro region. Clim. Chang. 2016, 138, 239–251. [Google Scholar] [CrossRef]
- Fraga, H.; García de Cortázar Atauri, I.; Santos, J.A. Viticultural irrigation demands under climate change scenarios in Portugal. Agric. Water Manag. 2018, 196, 66–74. [Google Scholar] [CrossRef]
- Costa, J.M.; Vaz, M.; Escalona, J.; Egipto, R.; Lopes, C.; Medrano, H.; Chaves, M.M. Modern viticulture in southern Europe: Vulnerabilities and strategies for adaptation to water scarcity. Agric. Water Manag. 2016, 164, 5–18. [Google Scholar] [CrossRef]
- Brito, C.; Dinis, L.T.; Moutinho-Pereira, J.; Correia, C. Kaolin, an emerging tool to alleviate the effects of abiotic stresses on crop performance. Sci. Hortic. (Amst.) 2019, 250, 310–316. [Google Scholar] [CrossRef]
- Dinis, L.T.; Ferreira, H.; Pinto, G.; Bernardo, S.; Correia, C.M.; Moutinho-Pereira, J. Kaolin-based, foliar reflective film protects photosystem II structure and function in grapevine leaves exposed to heat and high solar radiation. Photosynthetica 2016, 54, 47–55. [Google Scholar] [CrossRef]
- Dinis, L.T.; Malheiro, A.C.; Luzio, A.; Fraga, H.; Ferreira, H.; Gonçalves, I.; Pinto, G.; Correia, C.M.; Moutinho-Pereira, J. Improvement of grapevine physiology and yield under summer stress by kaolin-foliar application: Water relations, photosynthesis and oxidative damage. Photosynthetica 2018, 56, 641–651. [Google Scholar] [CrossRef]
- Conde, A.; Neves, A.; Breia, R.; Pimentel, D.; Dinis, L.T.; Bernardo, S.; Correia, C.M.; Cunha, A.; Gerós, H.; Moutinho-Pereira, J. Kaolin particle film application stimulates photoassimilate synthesis and modifies the primary metabolome of grape leaves. J. Plant Physiol. 2018, 223, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Dinis, L.T.; Bernardo, S.; Conde, A.; Pimentel, D.; Ferreira, H.; Félix, L.; Gerós, H.; Correia, C.M.; Moutinho-Pereira, J. Kaolin exogenous application boosts antioxidant capacity and phenolic content in berries and leaves of grapevine under summer stress. J. Plant Physiol. 2016, 191, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Conde, A.; Pimentel, D.; Neves, A.; Dinis, L.-T.; Bernardo, S.; Correia, C.M.; Gerós, H.; Moutinho-Pereira, J. Kaolin Foliar Application Has a Stimulatory Effect on Phenylpropanoid and Flavonoid Pathways in Grape Berries. Front. Plant Sci. 2016, 7, 1–14. [Google Scholar] [CrossRef]
- Poni, S.; Gatti, M.; Palliotti, A.; Dai, Z.; Duchêne, E.; Truong, T.T.; Ferrara, G.; Matarrese, A.M.S.; Gallotta, A.; Bellincontro, A.; et al. Grapevine quality: A multiple choice issue. Sci. Hortic. (Amst.) 2018, 234, 445–462. [Google Scholar] [CrossRef] [Green Version]
- Breia, R.; Vieira, S.; Da Silva, J.M.; Gerós, H.; Cunha, A. Mapping grape berry photosynthesis by chlorophyll fluorescence imaging: The effect of saturating pulse intensity in different tissues. Photochem. Photobiol. 2013, 89, 579–585. [Google Scholar] [CrossRef]
- Garrido, A.; Breia, R.; Serôdio, J.; Cunha, A. Impact of the Light Microclimate on Photosynthetic Activity of Grape Berry (Vitis vinifera): Insights for Radiation Absorption Mitigations’ Measures. In Theory and Practice of Climate Adaptation; Springer: Cham, Switzerland, 2018; pp. 419–441. [Google Scholar]
- Le Grange, M.; Wand, S.J.E.; Theron, K.I. Effect of kaolin applications on apple fruit quality and gas exchange of apple leaves. Acta Hortic. 2004, 636, 545–550. [Google Scholar] [CrossRef]
- Wünsche, J.-N.; Lombardini, L.; Greer, D.H.; Palmer, J.W. “Surround” particle film applications—The effect on whole canopy physiology of apple. Acta Hortic. 2004, 636, 565–571. [Google Scholar] [CrossRef]
- Rosati, A.; Metcalf, S.G.; Buchner, R.P.; Fulton, A.E.; Lampinen, B.D. Physiological effects of kaolin applications in well-irrigated and water-stressed walnut and almond trees. Ann. Bot. 2006, 98, 267–275. [Google Scholar] [CrossRef] [PubMed]
- Rosati, A.; Metcalf, S.G.; Buchner, R.P.; Fulton, A.E.; Lampinen, B.D. Effects of Kaolin application on light absorption and distribution, radiation use efficiency and photosynthesis of almond and walnut canopies. Ann. Bot. 2007, 99, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Glenn, D.M.; Puterka, G.J. The use of plastic films and sprayable reflective particle films to increase light penetration in apple canopies and improve apple color and weight. HortScience 2007, 42, 91–96. [Google Scholar] [CrossRef]
- Glenn, D.M. Particle Film Mechanisms of Action That Reduce the Effect of Environmental Stress in ‘Empire’ Apple. J. Am. Soc. Hortic. Sci. 2009, 134, 314–321. [Google Scholar] [CrossRef]
- Escalona, J.M.; Flexas, J.; Bota, J.; Medrano, H. Distribution of leaf photosynthesis and transpiration within grapevine canopies under different drought conditions. Vitis 2003, 42, 57–64. [Google Scholar]
- Obiadalla-Ali, H.; Fernie, A.R.; Lytovchenko, A.; Kossmann, J.; Lloyd, J.R. Inhibition of chloroplastic fructose 1,6-bisphosphatase in tomato fruits leads to decreased fruit size, but only small changes in carbohydrate metabolism. Planta 2004, 219, 533–540. [Google Scholar] [CrossRef]
- Cocaliadis, M.F.; Fernández-Muñoz, R.; Pons, C.; Orzaez, D.; Granell, A. Increasing tomato fruit quality by enhancing fruit chloroplast function. A double-edged sword? J. Exp. Bot. 2014, 65, 4589–4598. [Google Scholar] [CrossRef]
- Lorenz, D.H.; Eichhorn, K.W.; Bleiholder, H.; Klose, R.; Meier, U.; Weber, E. Growth Stages of the Grapevine: Phenological growth stages of the grapevine (Vitis vinifera L. ssp. vinifera)—Codes and descriptions according to the extended BBCH scale. Austr. J. Grape Wine Res. 1995, 1, 100–110. [Google Scholar] [CrossRef]
- Baker, N.R. Chlorophyll fluorescence: A probe of photosynthesis in vivo. Annu. Rev. Plant Biol. 2008, 59, 89–113. [Google Scholar] [CrossRef]
- Schreiber, U. Pulse-Amplitude-Modulation (PAM) Fluorometry and Saturation Pulse Method: An Overview. In Chlorophyll a Fluorescence: A Signature of Photosynthesis; Kluwer Academic: Dordrecht, The Netherlands, 2004; pp. 279–319. [Google Scholar]
- Genty, B.; Briantais, J.-M.; Baker, N.R. The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Biochim. Biophys. Acta Gen. Subj. 1989, 990, 87–92. [Google Scholar] [CrossRef]
- Fraser, P.D.; Pinto, M.E.S.; Holloway, D.E.; Bramley, P.M. Application of high-performance liquid chromatography with photodiode array detection to the metabolic profiling of plant isoprenoids. Plant J. 2000, 24, 551–558. [Google Scholar] [CrossRef] [PubMed]
- Mokochinski, J.B.; Mazzafera, P.; Sawaya, A.C.H.F.; Mumm, R.; de Vos, R.C.H.; Hall, R.D. Metabolic responses of Eucalyptus species to different temperature regimes. J. Integr. Plant Biol. 2018, 60, 397–411. [Google Scholar] [CrossRef]
- IPMA Instituto Português do Mar e da Atmosfera. Available online: http://www.ipma.pt/pt/otempo/obs.superficie/index-map-dia-chart.jsp#Braga, Merelim (accessed on 2 June 2019).
- IVV, Instituto da Vinha e do Vinho, I.P.I. de Mercado. Nota Informativa, No3/2018, 30/07/2018, Previsão de Colheita—Campanha 2018/2019. Available online: https://www.ivv.gov.pt/np4/8955.html (accessed on 1 July 2019).
- Shellie, K.C.; King, B.A. Kaolin particle film and water deficit influence malbec leaf and berry temperature, pigments, and photosynthesis. Am. J. Enol. Vitic. 2013, 64, 223–230. [Google Scholar] [CrossRef]
- Oliveira, M. Viticulture in Warmer Climates: Mitigating Environmental Stress in Douro Region, Portugal. In Grapes and Wines—Advances in Production, Processing, Analysis and Valorization; InTech: Rijeka, Croatia, 2018. [Google Scholar] [Green Version]
- Reynolds, A.G.; Heuvel, J.E.V. Influence of grapevine training systems on vine growth and fruit composition: A review. Am. J. Enol. Vitic. 2009, 60, 251–268. [Google Scholar]
- Kraus, C.; Pennington, T.; Herzog, K.; Hecht, A.; Fischer, M.; Voegele, R.T.; Hoffmann, C.; Töpfer, R.; Kicherer, A. Effects of canopy architecture and microclimate on grapevine health in two training systems. Vitis J. Grapevine Res. 2018, 57, 53–60. [Google Scholar] [CrossRef]
- Dos Santos, T.P.; Lopes, C.M.; Lucília Rodrigues, M.; de Souza, C.R.; Ricardo-da-Silva, J.M.; Maroco, J.P.; Pereira, J.S.; Manuela Chaves, M. Effects of deficit irrigation strategies on cluster microclimate for improving fruit composition of Moscatel field-grown grapevines. Sci. Hortic. (Amst.) 2007, 112, 321–330. [Google Scholar] [CrossRef]
- Sharma, R.R.; Vijay Rakesh Reddy, S.; Datta, S.C. Particle films and their applications in horticultural crops. Appl. Clay Sci. 2015, 116–117, 54–68. [Google Scholar] [CrossRef]
- Glenn, D.M.; Prado, E.; Erez, A.; McFerson, J.; Puterka, G.J. A Reflective, Processed-Kaolin Particle Film Affects Fruit Temperature, Radiation Reflection, and Solar Injury in Apple. J. Am. Soc. Hortic. Sci. 2002, 127, 188–193. [Google Scholar] [CrossRef] [Green Version]
- Glenn, M.D. The mechanisms of plant stress mitigation by kaolin-based particle films and applications in horticultural and agricultural crops. HortScience 2012, 47, 710–711. [Google Scholar] [CrossRef]
- Aschan, G.; Pfanz, H. Non-foliar photosynthesis—A strategy of additional carbon acquisition. Flora 2003, 198, 81–97. [Google Scholar] [CrossRef]
- Kadir, S.; Von Weihe, M.; Al-Khatib, K. Photochemical Efficiency and Recovery of Photosystem II in Grapes After Exposure to Sudden and Gradual Heat Stress. J. Am. Soc. Hortic. Sci. 2007, 132, 764–769. [Google Scholar] [CrossRef] [Green Version]
- Greer, D.H.; Weston, C. Heat stress affects flowering, berry growth, sugar accumulation and photosynthesis of Vitis vinifera cv. Semillon grapevines grown in a controlled environment. Funct. Plant Biol. 2010, 37, 206–214. [Google Scholar] [CrossRef]
- Luo, H.B.; Ma, L.; Xi, H.F.; Duan, W.; Li, S.H.; Loescher, W.; Wang, J.F.; Wang, L.J. Photosynthetic responses to heat treatments at different temperatures and following recovery in grapevine (Vitis amurensis L.) leaves. PLoS ONE 2011, 6. [Google Scholar] [CrossRef]
- Xiao, F.; Yang, Z.Q.; Lee, K.W. Photosynthetic and physiological responses to high temperature in grapevine (Vitis vinifera L.) leaves during the seedling stage. J. Hortic. Sci. Biotechnol. 2017, 92, 2–10. [Google Scholar] [CrossRef]
- Krause, G.H.; Jahns, P. Non-photochemical Energy Dissipation Determined by Chlorophyll Fluorescence Quenching: Characterization and Function. In Chlorophyll a Fluorescence; Springer: Dordrecht, The Netherlands, 2007. [Google Scholar]
- Medrano, H.; Bota, J.; Abadía, A.; Sampol, B.; Escalona, J.M.; Flexas, J. Effects of drought on light-energy dissipation mechanisms in high-light-acclimated, field-grown grapevines. Funct. Plant Biol. 2002, 29, 1197–1207. [Google Scholar] [CrossRef] [Green Version]
- Szabó, I.; Bergantino, E.; Giacometti, G.M. Light and oxygenic photosynthesis: Energy dissipation as a protection mechanism against photo-oxidation. EMBO Rep. 2005, 6, 629–634. [Google Scholar] [CrossRef]
- Joubert, C.; Young, P.R.; Eyéghé-Bickong, H.A.; Vivier, M.A. Field-Grown Grapevine Berries Use Carotenoids and the Associated Xanthophyll Cycles to Acclimate to UV Exposure Differentially in High and Low Light (Shade) Conditions. Front. Plant Sci. 2016, 7, 1–17. [Google Scholar] [CrossRef]
- Tikkanen, M.; Grieco, M.; Nurmi, M.; Rantala, M.; Suorsa, M.; Aro, E.M. Regulation of the photosynthetic apparatus under fluctuating growth light. Philos. Trans. R. Soc. B Biol. Sci. 2012, 367, 3486–3493. [Google Scholar] [CrossRef] [Green Version]
- Frioni, T.; Tombesi, S.; Luciani, E.; Sabbatini, P.; Berrios, J.G.; Palliotti, A. Kaolin treatments on Pinot noir grapevines for the control of heat stress damages. Bio Web Conf. 2019, 13, 04004. [Google Scholar] [CrossRef]
- Brito, C.; Dinis, L.T.; Luzio, A.; Silva, E.; Gonçalves, A.; Meijón, M.; Escandón, M.; Arrobas, M.; Rodrigues, M.Â.; Moutinho-Pereira, J.; et al. Kaolin and salicylic acid alleviate summer stress in rainfed olive orchards by modulation of distinct physiological and biochemical responses. Sci. Hortic. (Amst.) 2019, 246, 201–211. [Google Scholar] [CrossRef]
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence—A practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Cooley, N.M.; Glenn, D.M.; Clingeleffer, P.R.; Walker, R.R. The Effects of Water Deficit and Particle Film Technology Interactions on Cabernet Sauvignon Grape Composition. Acta Hortic. 2008, 193–200. [Google Scholar] [CrossRef]
- Shellie, K.; Glenn, D.M. Wine grape response to foliar kaolin particle film under differing levels of preveraison water stress. HortScience 2008, 43, 1392–1397. [Google Scholar] [CrossRef]
- Glenn, M.; Cooley, N.; Walker, R.; Clingeleffer, P.; Shellie, K. Impact of kaolin particle film and water deficit on wine grape water use efficiency and plant water relations. HortScience 2010, 45, 1178–1187. [Google Scholar] [CrossRef]
- Shellie, K.C.; King, B.A. Kaolin particle film and water deficit influence red winegrape color under high solar radiation in an arid climate. Am. J. Enol. Vitic. 2013, 64, 214–222. [Google Scholar] [CrossRef]
Figure 1.
(A) Timeline of the grape growing season depicting the sampling times, foliar kaolin application dates, and the onset of irrigation. (B) Scheme of treatment combinations applied in the field: irrigation (I)/ non-irrigation (NI) × kaolin (K)/ non-kaolin (NK). (WAA - weeks after anthesis).
Figure 1.
(A) Timeline of the grape growing season depicting the sampling times, foliar kaolin application dates, and the onset of irrigation. (B) Scheme of treatment combinations applied in the field: irrigation (I)/ non-irrigation (NI) × kaolin (K)/ non-kaolin (NK). (WAA - weeks after anthesis).

Figure 2.
Light intensities received by LL and HL clusters at the green stage (a,b) and the mature stage (c,d), for plants with irrigation (blue columns, note: the textured blue columns at green stage i.e., before the onset of irrigation, represent the measurements in the plots that were later irrigated) and foliar kaolin application (white columns). Black columns correspond to the respective controls. Values represent means with a standard deviation (n = 16 plants). Statistical notation: per ripening stage, different capital letters refer to significant differences (two-way ANOVA, p ≤ 0.05) between the two light microclimates within the same plant treatment, and different lowercase letters for differences between treatments within each light microclimate. If the respective factor did not have a significant effect, the lowercase letters were omitted.
Figure 2.
Light intensities received by LL and HL clusters at the green stage (a,b) and the mature stage (c,d), for plants with irrigation (blue columns, note: the textured blue columns at green stage i.e., before the onset of irrigation, represent the measurements in the plots that were later irrigated) and foliar kaolin application (white columns). Black columns correspond to the respective controls. Values represent means with a standard deviation (n = 16 plants). Statistical notation: per ripening stage, different capital letters refer to significant differences (two-way ANOVA, p ≤ 0.05) between the two light microclimates within the same plant treatment, and different lowercase letters for differences between treatments within each light microclimate. If the respective factor did not have a significant effect, the lowercase letters were omitted.

Figure 3.
Temperatures of LL and HL clusters at the green stage (a,b) and the mature stage (c,d), for plants with irrigation (blue columns, note: the textured blue columns at the green stage i.e., before the onset of irrigation, represent the measurements in the plots that were later irrigated) and foliar kaolin application (white columns). Black columns correspond to the respective controls. Values represent means with standard deviation (n = 16 plants). Statistical notation: per ripening stage, different capital letters refer to significant differences (two-way ANOVA, p ≤ 0.05) between the two light microclimates within the same plant treatment, and different lowercase letters for differences between treatments within each light microclimate. If the respective factor did not have a significant effect, the lowercase letters were omitted.
Figure 3.
Temperatures of LL and HL clusters at the green stage (a,b) and the mature stage (c,d), for plants with irrigation (blue columns, note: the textured blue columns at the green stage i.e., before the onset of irrigation, represent the measurements in the plots that were later irrigated) and foliar kaolin application (white columns). Black columns correspond to the respective controls. Values represent means with standard deviation (n = 16 plants). Statistical notation: per ripening stage, different capital letters refer to significant differences (two-way ANOVA, p ≤ 0.05) between the two light microclimates within the same plant treatment, and different lowercase letters for differences between treatments within each light microclimate. If the respective factor did not have a significant effect, the lowercase letters were omitted.

Figure 4.
Transmittance (%) spectrum (a) of a kaolin suspension (5% w/v) and reflectance (%) spectra (b) of leaves with and without kaolin (control) (n = 3).
Figure 4.
Transmittance (%) spectrum (a) of a kaolin suspension (5% w/v) and reflectance (%) spectra (b) of leaves with and without kaolin (control) (n = 3).

Figure 5.
Maximum quantum efficiency of PSII (Fv/Fm) mean values (n = 12–24 berries, +SD) of exocarps and seed integuments obtained from dark-adapted LL and HL grape berries grown under the four combinations of the two treatments applied: irrigation (I)/ non-irrigation (NI) × kaolin (K)/ non-kaolin (NK). Samples were collected at three development stages (green, véraison, and mature). Statistical notation: for each developmental stage, capital letters refer to differences between light microclimates within the same treatment combination, and lowercase letters refer to differences between treatment combinations within each light microclimate (mean values with a common letter were not significantly different). When capital and lowercase letters are omitted, the respective factor did not have a significant effect (two-way ANOVA p > 0.05). Notation with an asterisk means that only one factor (kaolin or irrigation) was significant.
Figure 5.
Maximum quantum efficiency of PSII (Fv/Fm) mean values (n = 12–24 berries, +SD) of exocarps and seed integuments obtained from dark-adapted LL and HL grape berries grown under the four combinations of the two treatments applied: irrigation (I)/ non-irrigation (NI) × kaolin (K)/ non-kaolin (NK). Samples were collected at three development stages (green, véraison, and mature). Statistical notation: for each developmental stage, capital letters refer to differences between light microclimates within the same treatment combination, and lowercase letters refer to differences between treatment combinations within each light microclimate (mean values with a common letter were not significantly different). When capital and lowercase letters are omitted, the respective factor did not have a significant effect (two-way ANOVA p > 0.05). Notation with an asterisk means that only one factor (kaolin or irrigation) was significant.

Figure 6.
Mean values (n = 12–24 berries, +SD) of relative rate of electron transport through PSII at 200 µmol photons m−2 s−1 (rETR200). All the microclimate conditions, treatment combinations and statistical information are the same as in Figure 5.
Figure 6.
Mean values (n = 12–24 berries, +SD) of relative rate of electron transport through PSII at 200 µmol photons m−2 s−1 (rETR200). All the microclimate conditions, treatment combinations and statistical information are the same as in Figure 5.

Figure 7.
Non-photochemical quenching (NPQ) mean values (n = 12–24 berries, +SD). All the microclimate conditions, treatment combinations, and statistical information are the same as in Figure 5.
Figure 7.
Non-photochemical quenching (NPQ) mean values (n = 12–24 berries, +SD). All the microclimate conditions, treatment combinations, and statistical information are the same as in Figure 5.

Figure 8.
Chlorophylls (a,b) and carotenoids (c,d) concentration mean values (n = 3, +SD) of exocarps and seeds obtained from LL and HL grape berries grown under non-kaolin (black columns) and kaolin (white columns) application, and collected at the green stage. Statistical notation: different capital letters refer to significant differences (two-way ANOVA, p ≤ 0.05) between the two light microclimates within the same plant treatment, and different lowercase letters to differences between treatments within each light microclimate. If the respective factor did not have a significant effect, the letters were omitted.
Figure 8.
Chlorophylls (a,b) and carotenoids (c,d) concentration mean values (n = 3, +SD) of exocarps and seeds obtained from LL and HL grape berries grown under non-kaolin (black columns) and kaolin (white columns) application, and collected at the green stage. Statistical notation: different capital letters refer to significant differences (two-way ANOVA, p ≤ 0.05) between the two light microclimates within the same plant treatment, and different lowercase letters to differences between treatments within each light microclimate. If the respective factor did not have a significant effect, the letters were omitted.

© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Garrido, A.; Serôdio, J.; De Vos, R.; Conde, A.; Cunha, A. Influence of Foliar Kaolin Application and Irrigation on Photosynthetic Activity of Grape Berries. Agronomy 2019, 9, 685. https://doi.org/10.3390/agronomy9110685
AMA Style
Garrido A, Serôdio J, De Vos R, Conde A, Cunha A. Influence of Foliar Kaolin Application and Irrigation on Photosynthetic Activity of Grape Berries. Agronomy. 2019; 9(11):685. https://doi.org/10.3390/agronomy9110685
Chicago/Turabian StyleGarrido, Andreia, João Serôdio, Ric De Vos, Artur Conde, and Ana Cunha. 2019. "Influence of Foliar Kaolin Application and Irrigation on Photosynthetic Activity of Grape Berries" Agronomy 9, no. 11: 685. https://doi.org/10.3390/agronomy9110685
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.