Genomic Regions Associated with IgE Levels against Culicoides spp. Antigens in Three Horse Breeds


 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Phenotype–IgE Levels Determined with ELISA Test
2.3. DNA Extractions, Genotyping, and Quality Control
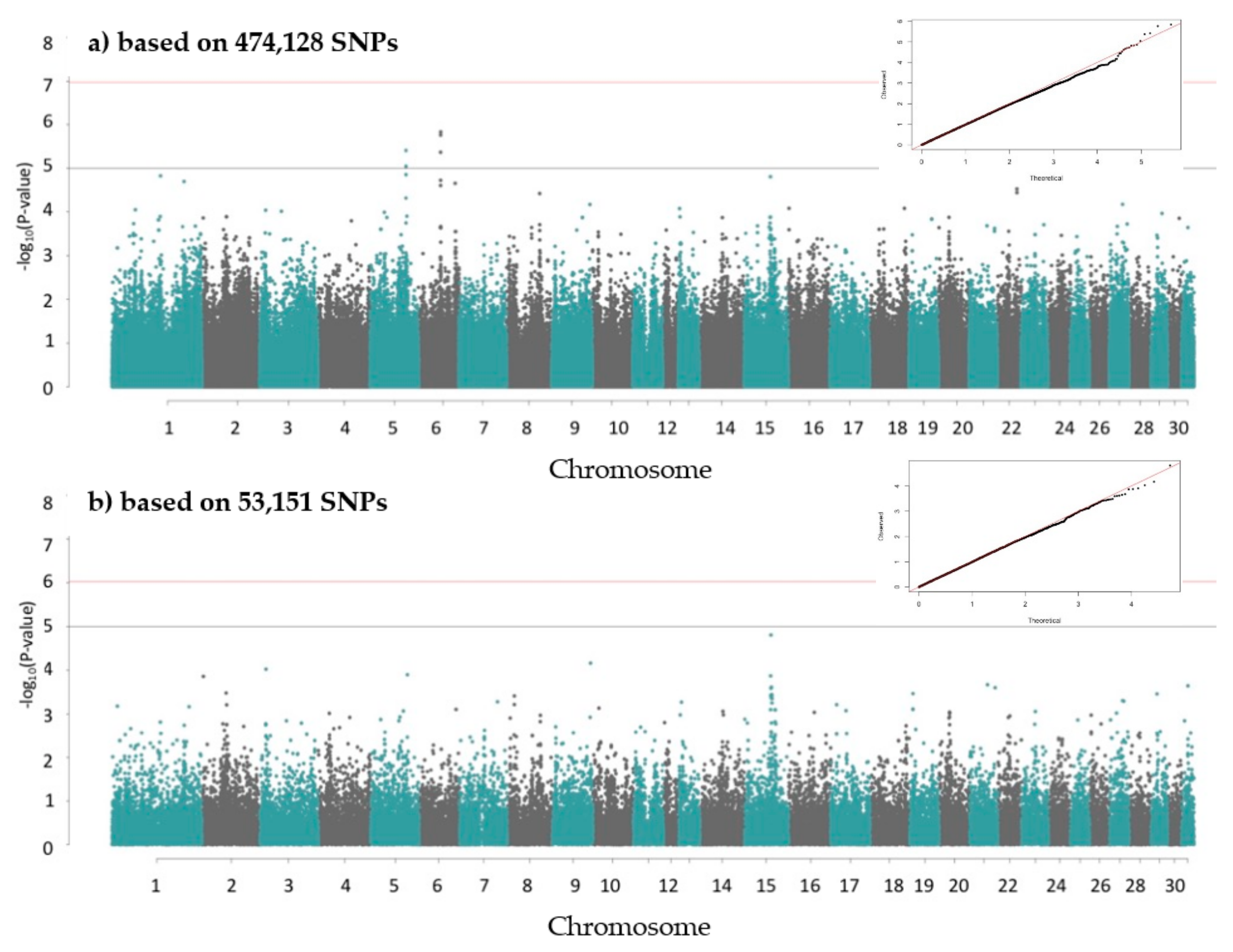
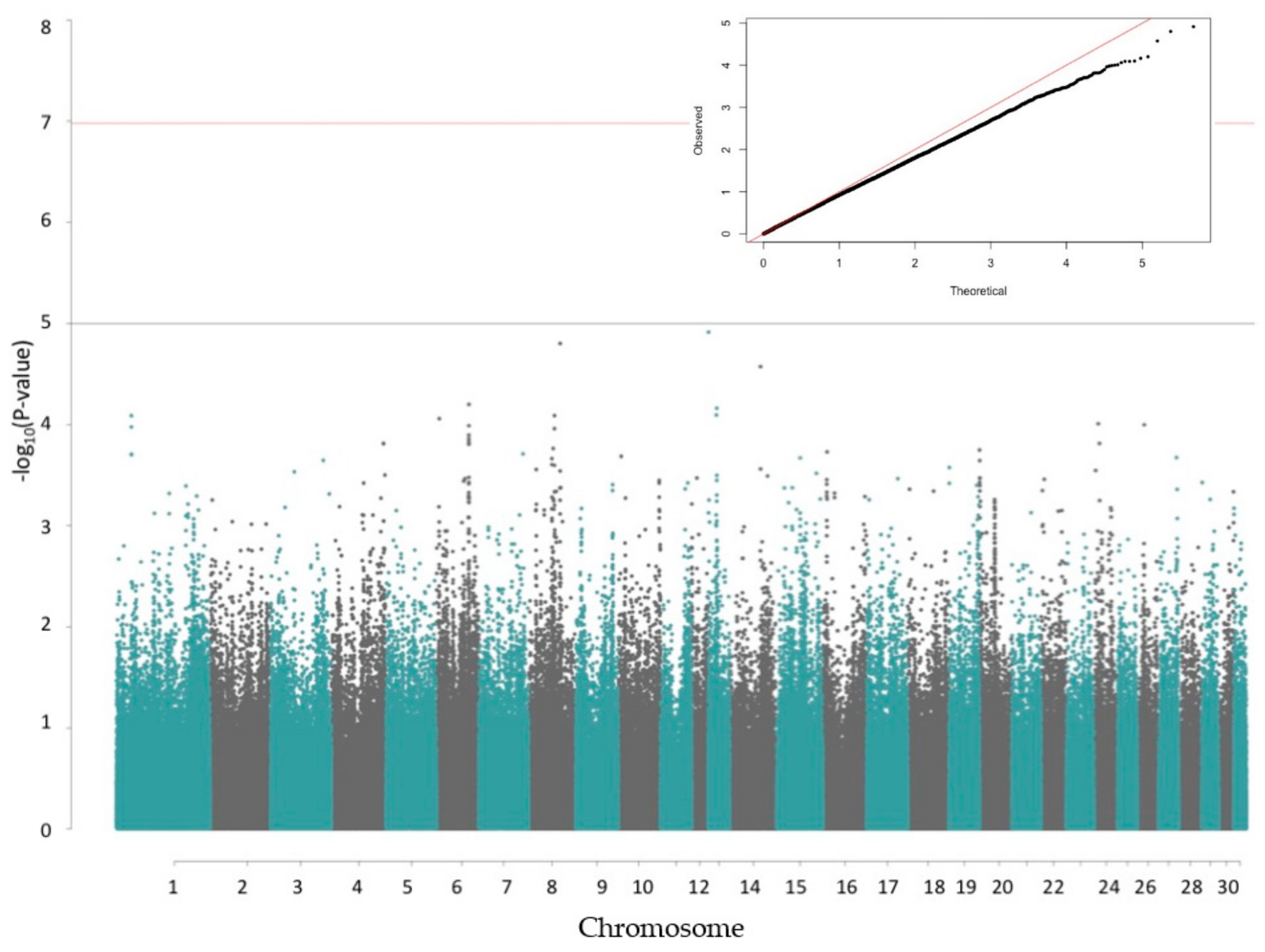
2.4. Genome-Wide Association (GWA) Study
3. Results
4. Discussions
5. Conclusions and Implications
Author Contributions
Funding
Conflicts of Interest
Appendix A

Appendix B

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Breed | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Belgian Warmblood Horse | Icelandic Horse | Shetland Pony | |||||||
| OD450nm Values Mean (SD) Minimum Maximum | Cases | Controls | Total | Cases | Controls | Total | Cases | Controls | Total |
| Culo1a 2 | 0.52 (0.78) 0.001 3.64 | 0.07 (0.11) 0.003 0.64 | 0.31 (0.62) 0.0003 3.64 | − | − | − | − | − | − |
| Culo1b 2 | − | − | − | 0.89 (0.43) 0.08 2.18 | 0.18 (0.27) 0.06 1.80 | 0.50 (0.38) 0.06 2.18 | 0.80 (0.44) 0.08 2.21 | 0.22 (0.12) 0.08 1.02 | 0.51 (0.35) 0.08 2.21 |
| Culo2a 2 | 0.53 (0.91) 0.0007 3.43 | 0.08 (0.09) 0.0005 0.43 | 0.33 (0.72) 0.0005 3.43 | − | − | − | − | − | − |
| Culo2b 2 | − | − | − | 0.42 (0.61) 0.12 2.18 | 0.15 (0.24) 0.07 1.41 | 0.27 (0.61) 0.07 2.18 | 0.44 (0.62) 0.16 2.12 | 0.17 (0.35) 0.08 2.23 | 0.31 (0.64) 0.08 2.23 |
| Culo3 | − | − | − | 1.07 (0.63) 0.13 2.67 | 0.22 (0.32) 0.07 1.95 | 0.59 (0.64) 0.07 2.67 | 1.17 (0.61) 0.10 2.31 | 0.38 (0.40) 0.07 2.04 | 0.78 (0.65) 0.07 2.31 |
| Culo4 | − | − | − | 1.12 (0.37) 0.08 2.06 | 0.25 (0.28) 0.07 1.81 | 0.64 (0.34) 0.07 2.06 | 1.10 (0.44) 0.08 2.07 | 0.32 (0.22) 0.07 1.39 | 0.72 (0.38) 0.07 2.07 |
| Culo5 | − | − | − | 0.37 (0.56) 0.15 2.18 | 0.15 (0.28) 0.10 1.35 | 0.24 (0.56) 0.10 2.18 | 0.48 (0.59) 0.12 2.54 | 0.20 (0.51) 0.11 2.51 | 0.34 (0.68) 0.11 2.54 |
| Culo6 | − | − | − | 0.98 (0.37) 0.07 1.79 | 0.26 (0.20) 0.07 1.61 | 0.58 (0.31) 0.07 1.79 | 1.27 (0.45) 0.07 3.13 | 0.46 (0.15) 0.07 0.88 | 0.88 (0.37) 0.07 3.13 |
| Culo7 | − | − | − | 0.39 (0.56) 0.08 2.26 | 0.15 (0.43) 0.06 2.81 | 0.25 (0.51) 0.06 2.81 | 0.52 (0.61) 0.11 2.62 | 0.20 (0.40) 0.06 3.04 | 0.36 (0.55) 0.06 3.08 |
| Combi1 3 | − | − | − | 1.35 (0.58) 0.28 2.65 | 0.28 (0.37) 0.07 1.95 | 0.75 (0.71) 0.07 2.65 | 1.50 (0.61) 0.11 2.62 | 0.35 (0.45) 0.08 2.03 | 0.94 (0.79) 0.08 2.62 |
| Combi2 3 | − | − | − | 1.24 (0.62) 0.18 2.29 | 0.31 (0.43) 0.08 1.88 | 0.72 (0.70) 0.08 2.29 | 1.44 (0.63) 0.17 2.42 | 0.41 (0.48) 0.11 2.14 | 0.93 (0.76) 0.11 2.42 |
| Culn1 4 | 0.35 (0.73) 0.001 3.56 | 0.07 (0.12) 0.0003 0.65 | 0.23 (0.58) 0.0002 3.56 | − | − | − | − | − | − |
| Culn3 4 | 0.21 (0.40) 0.0003 2.90 | 0.09 (0.15) 0.0003 1.21 | 0.15 (0.31) 0.0002 2.90 | − | − | − | − | − | − |
| Culn4 4 | 0.35 (0.50) 0.002 2.83 | 0.11 (0.22) 0.0003 1.65 | 0.24 (0.42) 0.0002 2.83 | − | − | − | − | − | − |
| Culn5 4 | 0.17 (0.38) 0.001 2.94 | 0.04 (0.05) 0.0003 0.29 | 0.11 (0.29) 0.0003 2.95 | − | − | − | − | − | − |
| Culn7 4 | 0.15 (0.33) 0.005 2.92 | 0.07 (0.06) 0.0004 0.24 | 0.12 (0.25) 0.0005 2.92 | − | − | − | − | − | − |
| Culn8 4 | 0.05 (0.06) 0.0004 0.31 | 0.04 (0.04) 0.0007 0.22 | 0.05 (0.05) 0.0004 0.31 | − | − | − | − | − | − |
| Culn10 4 | 0.13 (0.17) 0.0007 0.84 | 0.09 (0.12) 0.0003 0.83 | 0.11 (0.15) 0.0003 0.84 | − | − | − | − | − | − |
| WBE 5 | 0.20 (0.20) 0.0002 1.04 | 0.09 (0.10) 0.0003 0.67 | 0.15 (0.17) 0.0002 1.04 | − | − | − | − | − | − |
| WBE 6 | − | − | − | 0.89 (0.49) 0.13 2.08 | 0.18 (0.21) 0.08 1.71 | 0.50 (0.51) 0.08 2.08 | 0.80 (0.43) 0.13 1.92 | 0.22 (0.12) 0.09 0.59 | 0.51 (0.44) 0.09 1.92 |
| nubTH 7 | 0.18 (0.18) 0.001 1.02 | 0.07 (0.08) 0.0006 0.35 | 0.14 (0.16) 0.0006 1.02 | − | − | − | − | − | − |
| Log transformed OD450nm values 1 Mean (SD) Minimum−maximum | |||||||||
| L_Culo1a 2 | 1.77 (1.64) −1.29 6.55 | 3.44 (1.42) 0.44 8.27 | 2.54 (1.75) −1.29 8.27 | − | − | − | − | − | − |
| L_Culo1b 2 | − | − | − | −1.27 (0.86) −2.47 0.78 | −2.27 (0.56) −2.77 0.59 | −1.83 (0.87) −2.77 0.78 | −1.19 (0.81) −2.54 0.79 | −1.90 (0.43) −2.58 0.02 | −1.54 (0.74) −2.58 0.79 |
| L_Culo2a 2 | 1.92 (1.80) −1.23 7.19 | 3.29 (1.41) 0.83 7.60 | 2.51 (1.77) −1.23 7.60 | − | − | − | − | − | − |
| L_Culo2b 2 | − | − | − | −0.19 (0.80) −2.15 0.78 | −1.76 (0.61) −2.65 0.34 | −1.07 (1.05) −2.65 0.78 | −0.05 (0.73) −1.85 0.75 | −1.26 (0.70) −2.56 0.80 | −0.64 (0.93) −2.56 0.80 |
| L_Culo3 | − | − | − | −0.13 (0.79) −2.07 0.98 | −1.76 (0.77) −2.72 0.67 | −1.04 (1.12) −2.72 0.98 | −0.14 (0.79) −2.35 0.84 | −1.57 (0.78) −2.71 0.71 | 0.84 (1.06) −2.71 0.84 |
| L_Culo4 | − | − | − | −1.32 (0.73) −2.47 0.72 | −2.25 (0.55) −2.70 0.59 | −1.84 (0.79) −2.70 0.72 | −1.08 (0.82) −2.52 0.73 | −1.89 (0.60) −2.64 0.33 | −1.48 (0.82) −2.64 0.73 |
| L_Culo5 | − | − | − | −0.23 (0.72) −1.90 0.78 | −1.64 (0.67) −2.28 0.30 | −1.02 (0.98) −2.28 0.78 | 0.07 (0.66) −2.13 0.93 | −1.12 (0.75) −2.21 0.92 | −0.51 (0.92) −2.21 0.93 |
| L_Culo6 | − | − | − | −1.29 (0.80) −2.60 0.58 | −2.17 (0.56) −2.72 0.47 | −1.78 (0.81) −2.72 0.58 | −0.96 (0.79) −2.64 1.14 | −1.79 (0.55) −2.61 −0.13 | −1.37 (0.80) −2.64 1.14 |
| L_Culo7 | − | − | − | −1.19 (0.98) −2.58 0.81 | −2.11 (0.78) −2.74 1.03 | −1.71 (0.99) −2.74 1.03 | −1.15 (0.99) −2.73 1.12 | −2.17 (0.56) −2.88 1.11 | −1.65 (0.96) −2.88 1.12 |
| L_Combi1 3 | − | − | − | 0.19 (0.51) −1.28 0.97 | −1.74 (0.83) −2.60 0.67 | −0.89 (1.19) −2.60 0.97 | 0.26 (0.64) −2.16 0.96 | −1.48 (0.79) −2.58 0.71 | −0.59 (1.13) −2.58 0.96 |
| L_Combi2 3 | − | − | − | 0.05 (0.62) −1.69 0.83 | −2.66 (0.82) −2.49 0.63 | −0.90 (1.12) −2.49 0.83 | 0.20 (0.66) −1.76 0.88 | −1.28 (0.77) −2.23 0.76 | −0.53 (1.03) −2.23 0.88 |
| L_Culn1 4 | 2.72 (1.89) −1.27 6.80 | 3.41 (1.44) 0.43 8.24 | 3.01 (1.75) −1.27 8.24 | − | − | − | − | − | − |
| L_Culn3 4 | 2.52 (1.51) −1.07 8.23 | 3.17 (1.35) −0.19 7.91 | 2.83 (1.47) −1.07 8.23 | − | − | − | − | − | − |
| L_Culn4 4 | 1.90 (1.48) −1.04 6.35 | 3.14 (1.49) −0.50 8.29 | 2.46 (1.61) −1.04 8.29 | − | − | − | − | − | − |
| L_Culn5 4 | 2.97 (1.66) −1.08 6.95 | 3.67 (1.27) 1.23 8.04 | 3.28 (1.53) −1.08 8.04 | − | − | − | − | − | − |
| L_Culn7 4 | 2.62 (1.20) −1.07 5.40 | 3.28 (1.29) 1.41 7.63 | 2.91 (1.28) −1.07 7.63 | − | − | − | − | − | − |
| L_Culn8 4 | 3.63 (1.44) 1.18 7.71 | 3.95 (1.32) 1.50 7.23 | 3.78 (1.39) 1.18 7.71 | − | − | − | − | − | − |
| L_Culn10 4 | 3.10 (1.76) 0.18 7.23 | 3.11 (1.42) 0.19 8.29 | 3.10 (1.62) 0.18 8.29 | − | − | − | − | − | − |
| L_WBE 5 | 2.28 (1.50) −0.04 8.34 | 3.02 (1.35) 0.40 8.27 | 2.61 (1.48) −0.04 8.34 | − | − | − | − | − | − |
| L_WBE 6 | − | − | − | −0.30 (0.67) −2.06 0.73 | −1.94 (0.53) −2.52 0.54 | −1.22 (1.01) −2.52 0.73 | −0.40 (0.64) −2.01 0.65 | −1.67 (0.49) −2.37 −0.52 | −1.02 (0.85) −2.37 0.65 |
| L_nubTH 7 | 2.19 (1.17) −0.02 6.70 | 3.53 (1.53) 1.06 7.40 | 2.72 (1.48) −1.23 7.60 | − | − | − | − | − | − |
Appendix C
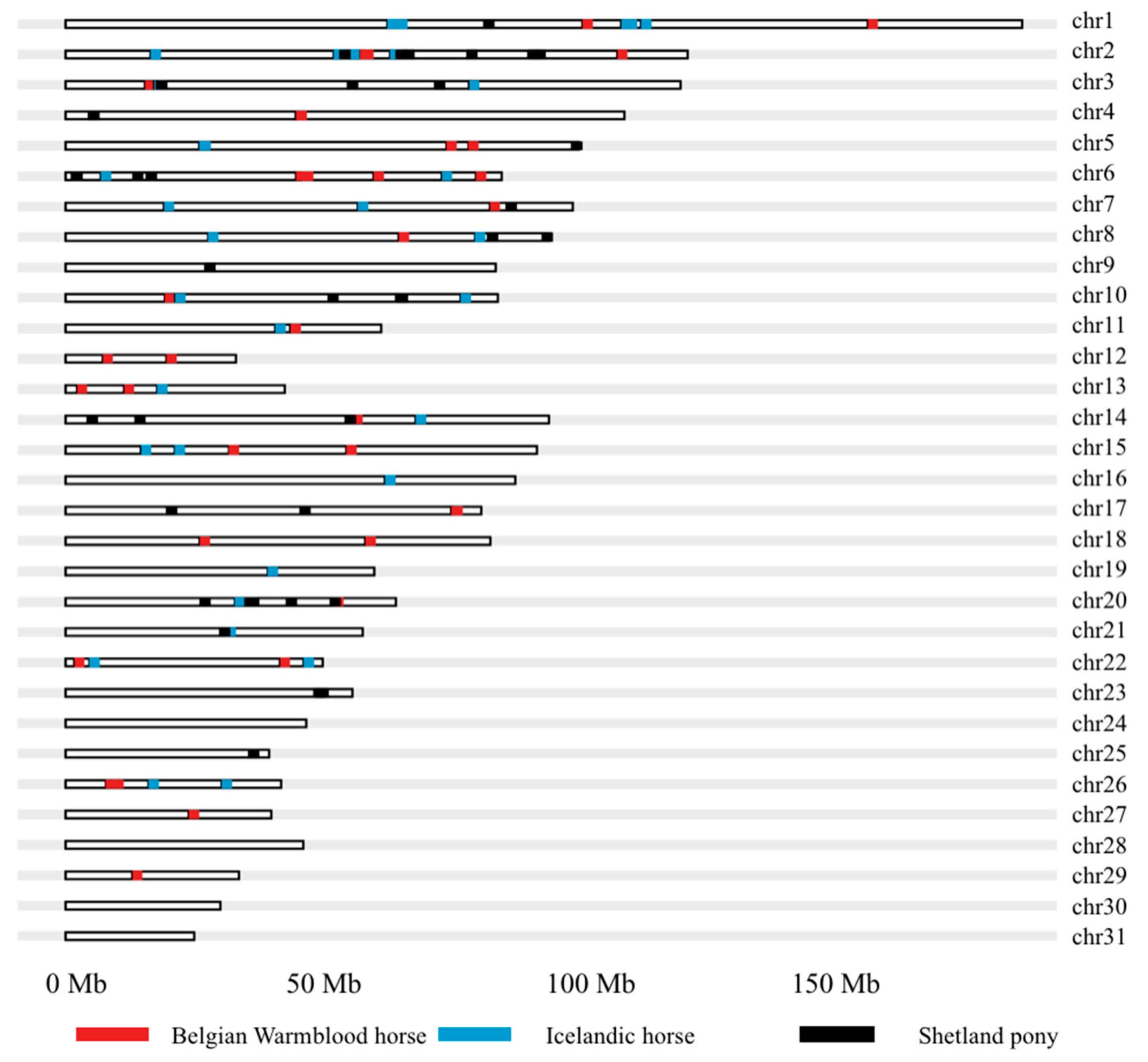
| SNP | Breed | Chr 1 | Pos 2 | IgE level 3 | A1 4 | A2 4 | Freq A2 5 | βSE 6 | Pc1df 7 |
|---|---|---|---|---|---|---|---|---|---|
| BIEC2_27204 | Icelandic horse | 1 | 62,517,648 | L_Culo6 | B | A | 0.03 | 1.3390.318 | 2.81 × 10−5 |
| BIEC2_27204 | Icelandic horse | 1 | 62,517,648 | L_Culo4 | B | A | 0.03 | 1.2130.308 | 1.06 × 10−4 |
| BIEC2_28234 | Icelandic horse | 1 | 64,386,608 | L_Culo6 | A | B | 0.05 | 0.9800.239 | 4.64 × 10−5 |
| BIEC2_37747 | Shetland pony | 1 | 81,341,895 | L_Culo4 | B | A | 0.14 | 0.4510.122 | 2.25 × 10−4 |
| AX-103503964 | Belgian Warmblood horse | 1 | 100,404,573 | L_Culn7 | A | G | 0.01 | 2.7660.639 | 1.50 × 10−5 |
| BIEC2_45214 | Icelandic horse | 1 | 107,931,611 | L_Culo4 | A | B | 0.21 | 0.4910.120 | 5.91 × 10−5 |
| BIEC2_45649 | Icelandic horse | 1 | 109,071,746 | L_Culo5 | B | A | 0.26 | 0.5690.144 | 8.00 × 10−5 |
| BIEC2_468481 | Icelandic horse | 1 | 111,803,287 | L_Culo4 | A | B | 0.08 | 0.8090.178 | 7.44 × 10−6 |
| BIEC2_468521 | Icelandic horse | 1 | 111,806,815 | L_Culo4 | A | B | 0.08 | 0.8090.178 | 7.44 × 10−6 |
| AX-103346536 | Belgian Warmblood horse | 1 | 155,846,866 | L_WBE | A | G | 0.02 | 2.7040.577 | 3.98 × 10−6 |
| BIEC2_458516 | Icelandic horse | 2 | 16,510,393 | L_Culo4 | B | A | 0.04 | 1.1120.272 | 5.09 × 10−5 |
| BIEC2_458516 | Icelandic horse | 2 | 16,510,393 | L_WBE | B | A | 0.04 | 1.3690.341 | 6.97 × 10−5 |
| BIEC2_477913 | Icelandic horse | 2 | 52,087,373 | L_Culo4 | B | A | 0.06 | 0.9350.232 | 7.27 × 10−5 |
| BIEC2_477926 | Icelandic horse | 2 | 52,126,031 | L_Culo4 | A | B | 0.06 | 0.9350.232 | 7.27 × 10−5 |
| BIEC2_477984 | Icelandic horse | 2 | 52,282,449 | L_Culo4 | B | A | 0.04 | 1.0530.257 | 5.59 × 10−5 |
| BIEC2_478173 | Shetland pony | 2 | 53,366,587 | L_Culo7 | B | A | 0.23 | 0.4400.111 | 9.34 × 10−5 |
| BIEC2-478967 | Icelandic horse | 2 | 55,097,850 | L_Culo6 | A | B | 0.01 | 2.1600.475 | 5.99 × 10−6 |
| BIEC2_478970 | Icelandic horse | 2 | 55,101,939 | L_Culo6 | B | A | 0.01 | 2.1600.475 | 5.99 × 10−6 |
| AX-104248441 | Belgian Warmblood horse | 2 | 57,180,618 | L_WBE | G | A | 0.30 | 0.7460.171 | 1.80 × 10−5 |
| AX-104413239 | Belgian Warmblood horse | 2 | 57,795,030 | L_WBE | G | A | 0.30 | 0.7590.168 | 8.58 × 10−6 |
| BIEC2_483351 | Icelandic horse | 2 | 63,034,801 | L_Culo1b | A | B | 0.04 | 1.1720.304 | 1.18 × 10−4 |
| BIEC2_484391 | Shetland pony | 2 | 64,289,972 | L_Culo7 | B | A | 0.04 | 0.9140.240 | 1.89 × 10−4 |
| BIEC2_484575 | Shetland pony | 2 | 65,851,405 | L_Culo7 | A | B | 0.04 | 0.9740.246 | 1.00 × 10−4 |
| BIEC2_491244 | Shetland pony | 2 | 78,068,427 | L_Culo6 | A | B | 0.37 | 0.3290.082 | 6.10 × 10−5 |
| BIEC2_522114 | Shetland pony | 2 | 89,890,319 | L_Culo7 | B | A | 0.21 | 0.4650.118 | 1.07 × 10−4 |
| TBIEC2_523156 | Shetland pony | 2 | 91,300,136 | L_Culo1b | B | A | 0.13 | 0.3890.104 | 1.81 × 10−4 |
| AX-104656854 | Belgian Warmblood horse | 2 | 107,184,258 | L_Culo1a | G | A | 0.01 | 3.8470.889 | 1.50 × 10−5 |
| AX-104651894 | Belgian Warmblood horse | 3 | 15,403,585 | L_Culn5 | C | T | 0.40 | −0.9190.210 | 1.78 × 10−5 |
| BIEC2_773441 | Icelandic horse | 3 | 17,214,985 | L_Combi1 | A | B | 0.12 | 0.9780.252 | 1.04 × 10−4 |
| BIEC2_813132 | Shetland pony | 3 | 17,751,670 | L_Culo1b | B | A | 0.10 | 0.4870.132 | 2.37 × 10−4 |
| BIEC2_782301 | Shetland pony | 3 | 54,836,579 | L_Culo3 | B | A | 0.47 | 0.4040.110 | 2.42 × 10−4 |
| BIEC2-791649 | Shetland pony | 3 | 71,803,910 | L_Culo4 | A | B | 0.02 | 1.3750.345 | 7.18 × 10−5 |
| BIEC2-791649 | Shetland pony | 3 | 71,803,910 | L_WBE | A | B | 0.02 | 1.4470.366 | 7.77 × 10−5 |
| BIEC2_794498 | Icelandic horse | 3 | 78,402,173 | L_Culo7 | B | A | 0.38 | 0.5320.131 | 5.15 × 10−5 |
| BIEC2_794498 | Icelandic horse | 3 | 78,402,173 | L_Culo5 | B | A | 0.38 | 0.5440.137 | 7.46 × 10−5 |
| BIEC2_794499 | Icelandic horse | 3 | 78,402,249 | L_Culo7 | A | B | 0.38 | 0.5320.131 | 5.15 × 10−5 |
| BIEC2_794499 | Icelandic horse | 3 | 78,402,249 | L_Culo5 | A | B | 0.38 | 0.5440.137 | 7.46 × 10−5 |
| BIEC2_844842 | Shetland pony | 4 | 4,540,728 | L_Culo6 | B | A | 0.06 | 0.7450.168 | 9.48 × 10−6 |
| BIEC2_844842 | Shetland pony | 4 | 4,540,728 | L_Culo4 | B | A | 0.06 | 0.7080.173 | 4.60 × 10−5 |
| BIEC2_844876 | Shetland pony | 4 | 4,590,809 | L_Culo6 | B | A | 0.09 | 0.5040.137 | 2.43 × 10−4 |
| AX-103506369 | Belgian Warmblood horse | 4 | 44,741,394 | L_Culn3 | T | C | 0.02 | −3.3380.755 | 1.00 × 10−5 |
| AX-103506369 | Belgian Warmblood horse | 4 | 44,741,394 | L_Culn1 | T | C | 0.02 | −3.3380.755 | 1.00 × 10−5 |
| AX-103506369 | Belgian Warmblood horse | 4 | 44,741,394 | L_Culn4 | T | C | 0.02 | −3.3380.755 | 1.00 × 10−5 |
| AX-103103078 | Belgian Warmblood horse | 4 | 44,929,049 | L_Culn3 | C | T | 0.02 | −3.3330.755 | 1.00 × 10−5 |
| AX-103103078 | Belgian Warmblood horse | 4 | 44,929,049 | L_Culn1 | C | T | 0.02 | −3.3330.755 | 1.00 × 10−5 |
| AX-103103078 | Belgian Warmblood horse | 4 | 44,929,049 | L_Culn4 | C | T | 0.02 | −3.3330.755 | 1.00 × 10−5 |
| BIEC2-898668 | Icelandic horse | 5 | 25,944,960 | L_Culo3 | B | A | 0.14 | 0.8420.214 | 8.50 × 10−5 |
| BIEC2_898714 | Icelandic horse | 5 | 26,215,264 | L_Culo5 | B | A | 0.35 | −0.5760.150 | 1.20 × 10−4 |
| AX-103271642 | Belgian Warmblood horse | 5 | 74,007,505 | L_Culn7 | A | G | 0.23 | −0.7250.163 | 9.08 × 10−6 |
| AX-104561711 | Belgian Warmblood horse | 5 | 74,007,642 | L_Culn7 | G | A | 0.23 | −0.7550.163 | 3.89 × 10−6 |
| AX-104085455 | Belgian Warmblood horse | 5 | 74,013,902 | L_Culn7 | C | T | 0.23 | −0.7120.164 | 1.41 × 10−5 |
| AX-103834157 | Belgian Warmblood horse | 5 | 78,269,939 | L_Culn5 | A | C | 0.30 | −1.0120.229 | 1.40 × 10−5 |
| BIEC2_933647 | Shetland pony | 5 | 98,392,335 | L_Culo7 | B | A | 0.03 | 1.1570.311 | 2.56 × 10−4 |
| BIEC2_935332 | Shetland pony | 6 | 1,212,183 | L_Culo4 | A | B | 0.10 | 0.5350.138 | 1.18 × 10−4 |
| BIEC2_983237 | Shetland pony | 6 | 1,328,824 | L_Culo4 | B | A | 0.10 | 0.5330.138 | 1.26 × 10−4 |
| BIEC2_938056 | Icelandic horse | 6 | 6,834,306 | L_Culo1b | A | B | 0.19 | 0.6320.140 | 6.24 × 10−6 |
| BIEC2-941245 | Shetland pony | 6 | 13,162,030 | L_Culo4 | B | A | 0.11 | 0.5740.134 | 2.10 × 10−5 |
| BIEC2_990454 | Shetland pony | 6 | 15,765,010 | L_Culo7 | B | A | 0.01 | 1.9490.437 | 1.18 × 10−5 |
| BIEC2_990454 | Shetland pony | 6 | 15,765,010 | L_Culo6 | B | A | 0.01 | 1.4050.367 | 1.27 × 10−4 |
| AX-104276231 | Belgian Warmblood horse | 6 | 44,759,822 | L_Culn8 | C | T | 0.06 | 1.8150.406 | 7.85 × 10−6 |
| AX-104807788 | Belgian Warmblood horse | 6 | 45,879,727 | L_Culn7 | A | G | 0.07 | 1.2840.279 | 4.27 × 10−6 |
| AX-103196232 | Belgian Warmblood horse | 6 | 45,975,297 | L_Culn7 | G | A | 0.10 | 1.0370.242 | 1.90 × 10−5 |
| AX-103555139 | Belgian Warmblood horse | 6 | 46,095,918 | L_Culn7 | T | C | 0.14 | 0.9970.208 | 1.72 × 10−6 |
| AX-103105514 | Belgian Warmblood horse | 6 | 46,125,756 | L_Culn7 | G | A | 0.14 | 1.0110.210 | 1.46 × 10−6 |
| AX-103811007 | Belgian Warmblood horse | 6 | 59,819,569 | L_WBE | T | C | 0.25 | 0.8400.193 | 1.78 × 10−5 |
| BIEC2_1017806 | Icelandic horse | 6 | 73,076,296 | L_Culo3 | B | A | 0.30 | 0.5710.140 | 4.63 × 10−5 |
| AX-103340585 | Belgian Warmblood horse | 6 | 79,707,409 | L_WBE | T | C | 0.01 | 2.7930.621 | 9.41 × 10−6 |
| BIEC2_984144 | Icelandic horse | 7 | 19,118,876 | L_Culo2b | A | B | 0.47 | 0.5350.133 | 5.64 × 10−5 |
| BIEC2_984146 | Icelandic horse | 7 | 19,118,997 | L_Culo2b | A | B | 0.47 | 0.5350.133 | 5.64 × 10−5 |
| BIEC2_1002598 | Icelandic horse | 7 | 56,761,597 | L_Culo1b | A | B | 0.40 | −0.4500.117 | 1.21 × 10−4 |
| AX-104003295 | Belgian Warmblood horse | 7 | 82,435,491 | L_WBE | T | C | 0.07 | 1.4690.305 | 2.09 × 10−6 |
| BIEC2-1010453 | Shetland pony | 7 | 85,717,480 | L_Culo6 | B | A | 0.55 | 0.2980.079 | 1.68 × 10−4 |
| BIEC2-1010454 | Shetland pony | 7 | 85,717,499 | L_Culo6 | A | B | 0.55 | 0.2980.079 | 1.68 × 10−4 |
| BIEC2_10104552 | Shetland pony | 7 | 85,717,606 | L_Culo6 | A | B | 0.55 | 0.2980.079 | 1.68 × 10−4 |
| BIEC2_1096840 | Icelandic horse | 8 | 27,964,452 | L_Culo7 | B | A | 0.07 | 1.0700.242 | 1.04 × 10−5 |
| AX-103533308 | Belgian Warmblood horse | 8 | 64,692,596 | L_WBE | A | G | 0.07 | 1.4050.314 | 1.08 × 10−5 |
| AX-103391816 | Belgian Warmblood horse | 8 | 64,784,788 | L_WBE | C | T | 0.06 | 1.4900.331 | 9.18 × 10−6 |
| BIEC2_1063702 | Icelandic horse | 8 | 79,554,471 | L_Culo4 | B | A | 0.02 | 1.3070.336 | 1.28 × 10−4 |
| BIEC2_1064164 | Shetland pony | 8 | 82,054,284 | L_Culo6 | B | A | 0.03 | −0.8430.230 | 2.41 × 10−4 |
| BIEC2_1125196 | Shetland pony | 8 | 92,699,636 | L_Culo4 | A | B | 0.09 | 0.5220.139 | 1.84 × 10−4 |
| BIEC2_1141819 | Shetland pony | 9 | 27,136,208 | L_Culo1b | B | A | 0.02 | 1.1030.282 | 8.98 × 10−5 |
| AX-104628650 | Belgian Warmblood horse | 10 | 19,284,909 | L_Culn5 | G | A | 0.08 | 1.6270.353 | 5.88 × 10−6 |
| BIEC2_108460 | Icelandic horse | 10 | 21,301,438 | L_Culo4 | B | A | 0.73 | 0.4530.113 | 8.24 × 10−5 |
| BIEC2_122837 | Shetland pony | 10 | 51,100,177 | L_Culo4 | A | B | 0.08 | 0.5590.153 | 2.68 × 10−4 |
| BIEC2_129582 | Shetland pony | 10 | 64,203,886 | L_Culo7 | B | A | 0.01 | 1.9110.435 | 1.57 × 10−5 |
| TBIEC2_136550 | Shetland pony | 10 | 64,532,326 | L_Culo7 | B | A | 0.26 | 0.4850.110 | 1.55 × 10−5 |
| BIEC2_131860 | Icelandic horse | 10 | 76,735,207 | L_Culo3 | B | A | 0.03 | 1.7840.451 | 7.54 × 10−5 |
| BIEC2_152809 | Icelandic horse | 11 | 40,721,405 | L_WBE | A | B | 0.47 | −0.5620.137 | 4.95 × 10−5 |
| AX-103887059 | Belgian Warmblood horse | 11 | 43,666,485 | L_Culn3 | C | T | 0.07 | −1.7950.415 | 1.55 × 10−5 |
| AX-103887059 | Belgian Warmblood horse | 11 | 43,666,485 | L_Culn1 | C | T | 0.07 | −1.7950.415 | 1.55 × 10−5 |
| AX-103887059 | Belgian Warmblood horse | 11 | 43,666,485 | L_Culn4 | C | T | 0.07 | −1.7950.415 | 1.55 × 10−5 |
| AX-103378324 | Belgian Warmblood horse | 12 | 7,216,566 | L_Culo2a | T | C | 0.02 | 3.1830.703 | 5.90 × 10−6 |
| AX-103538080 | Belgian Warmblood horse | 12 | 19,588,916 | L_Culo1a | T | G | 0.12 | −1.2000.278 | 1.60 × 10−5 |
| AX-104700154 | Belgian Warmblood horse | 13 | 2,214,862 | L_Culn8 | G | A | 0.48 | −0.8440.183 | 4.16 × 10−6 |
| AX-104619436 | Belgian Warmblood horse | 13 | 11,360,389 | L_WBE | C | T | 0.28 | 0.8030.172 | 4.18 × 10−6 |
| BIEC2_223584 | Icelandic horse | 13 | 17,801,684 | L_Culo5 | A | B | 0.30 | 0.5430.137 | 7.07 × 10−5 |
| BIEC2_223584 | Icelandic horse | 13 | 17,801,684 | L_Culo6 | A | B | 0.30 | 0.4580.116 | 9.00 × 10−5 |
| BIEC2-239709 | Shetland pony | 14 | 4,274,260 | L_Combi2 | B | A | 0.38 | 0.5270.136 | 1.03 × 10−4 |
| BIEC2_245596 | Shetland pony | 14 | 13,580,552 | L_Culo4 | A | B | 0.19 | −0.4200.104 | 5.28 × 10−5 |
| BIEC2_259420 | Shetland pony | 14 | 54,397,113 | L_Culo7 | B | A | 0.01 | 1.8730.482 | 1.33 × 10−4 |
| AX-104663195 | Belgian Warmblood horse | 14 | 55,773,239 | L_Culn8 | C | T | 0.02 | 3.0970.687 | 6.64 × 10−6 |
| BIEC2_262621 | Icelandic horse | 14 | 67,999,643 | L_Combi2 | B | A | 0.49 | −0.6480.158 | 3.99 × 10−5 |
| BIEC2_262673 | Icelandic horse | 14 | 68,068,555 | L_Combi2 | A | B | 0.53 | −0.6200.152 | 4.67 × 10−5 |
| BIEC2-289295 | Icelandic horse | 15 | 14,642,890 | L_Combi1 | B | A | 0.10 | −1.0040.241 | 3.00 × 10−5 |
| BIEC2-289295 | Icelandic horse | 15 | 14,642,890 | L_Combi2 | B | A | 0.10 | −0.9230.229 | 5.72 × 10−5 |
| BIEC2_289301 | Icelandic horse | 15 | 14,647,231 | L_Combi1 | A | B | 0.10 | −1.0040.241 | 3.00 × 10−5 |
| BIEC2_289301 | Icelandic horse | 15 | 14,647,231 | L_Combi2 | A | B | 0.10 | −0.9230.229 | 5.72 × 10−5 |
| BIEC2-294189 | Icelandic horse | 15 | 21,210,210 | L_Culo2b | B | A | 0.26 | 0.6510.191 | 5.23 × 10−5 |
| AX-103998137 | Belgian Warmblood horse | 15 | 31,741,601 | L_Culn8 | A | C | 0.44 | 0.7630.174 | 1.22 × 10−5 |
| AX-104442678 | Belgian Warmblood horse | 15 | 54,538,291 | L_Culn7 | G | A | 0.03 | −1.6410.380 | 1.57 × 10−5 |
| BIEC2_355030 | Icelandic horse | 16 | 62,046,387 | L_Combi2 | A | B | 0.19 | 0.7600.194 | 8.77 × 10−5 |
| BIEC2_355030 | Icelandic horse | 16 | 62,046,387 | L_Culo5 | A | B | 0.19 | 0.6390.166 | 1.21 × 10−4 |
| BIEC2_373511 | Shetland pony | 17 | 19,689,865 | L_Culo4 | B | A | 0.02 | 1.2290.319 | 1.21 × 10−4 |
| BIEC2_395885 | Shetland pony | 17 | 45,644,294 | L_Culo6 | B | A | 0.04 | −0.7440.203 | 2.42 × 10−4 |
| AX-103325608 | Belgian Warmblood horse | 17 | 74,894,091 | L_WBE | G | A | 0.01 | 3.8650.864 | 1.06 × 10−5 |
| AX-103932279 | Belgian Warmblood horse | 17 | 75,173,007 | L_WBE | T | C | 0.01 | 3.8650.864 | 1.06 × 10−5 |
| AX-104268962 | Belgian Warmblood horse | 17 | 75,184,430 | L_WBE | A | C | 0.02 | 3.4490.675 | 4.92 × 10−7 |
| AX-104514365 | Belgian Warmblood horse | 18 | 26,032,740 | L_Culn5 | A | G | 0.04 | 2.1700.499 | 1.90 × 10−5 |
| AX-103909004 | Belgian Warmblood horse | 18 | 58,215,227 | L_Culn10 | T | C | 0.01 | 4.2440.965 | 1.08 × 10−5 |
| AX-104652798 | Belgian Warmblood horse | 18 | 58,248,171 | L_Culn10 | G | T | 0.01 | 4.2500.965 | 1.05 × 10−5 |
| TBIEC2_461038 | Icelandic horse | 19 | 39,283,640 | L_Culo7 | B | A | 0.35 | −0.5290.132 | 6.43 × 10−5 |
| BIEC2_526197 | Shetland pony | 20 | 26,221,179 | L_Culo7 | B | A | 0.11 | 0.6790.158 | 2.31 × 10−5 |
| UKUL3411 | Icelandic horse | 20 | 32,902,939 | L_Culo4 | A | B | 0.16 | −0.6420.155 | 4.89 × 10−5 |
| BIEC2_558521 | Shetland pony | 20 | 34,924,555 | L_Culo7 | A | B | 0.59 | −0.3850.098 | 1.23 × 10−4 |
| UKUL3474 | Shetland pony | 20 | 35,643,200 | L_WBE | B | A | 0.53 | −0.4160.096 | 1.33 × 10−5 |
| UKUL3474 | Shetland pony | 20 | 35,643,200 | L_Culo7 | B | A | 0.53 | −0.3710.098 | 2.10 × 10−4 |
| UKUL3477 | Shetland pony | 20 | 35,692,652 | L_WBE | A | B | 0.45 | 0.4070.096 | 2.36 × 10−5 |
| UKUL3477 | Shetland pony | 20 | 35,692,652 | L_Culo7 | A | B | 0.45 | 0.3820.099 | 1.51 × 10−4 |
| BIEC2_532826 | Shetland pony | 20 | 42,983,718 | L_Culo1b | B | A | 0.45 | 0.2920.075 | 9.39 × 10−5 |
| BIEC2_538781 | Shetland pony | 20 | 51,509,431 | L_Culo1b | A | B | 0.27 | −0.3370.087 | 1.01 × 10−4 |
| AX-104089679 | Belgian Warmblood horse | 20 | 52,011,107 | L_nubTH | C | T | 0.02 | 3.7850.884 | 1.87 × 10−5 |
| BIEC2_560852 | Shetland pony | 21 | 30,022,509 | L_Culo7 | B | A | 0.37 | 0.3720.099 | 2.38 × 10−4 |
| BIEC2_560854 | Shetland pony | 21 | 30,022,801 | L_Culo7 | A | B | 0.37 | 0.3720.099 | 2.38 × 10−4 |
| BIEC2_560857 | Shetland pony | 21 | 30,033,646 | L_Culo7 | B | A | 0.37 | 0.3720.099 | 2.38 × 10−4 |
| BIEC2_561034 | Icelandic horse | 21 | 31,044,321 | L_Culo6 | A | B | 0.04 | 1.0690.272 | 8.97 × 10−5 |
| AX-103656067 | Belgian Warmblood horse | 22 | 1,680,310 | L_WBE | C | T | 0.11 | 1.2340.258 | 2.42 × 10−6 |
| BIEC2_577012 | Icelandic horse | 22 | 4,630,951 | L_Culo7 | A | B | 0.10 | 0.8450.213 | 7.31 × 10−5 |
| AX-104834793 | Belgian Warmblood horse | 22 | 41,631,523 | L_Culn5 | G | A | 0.04 | 2.2360.487 | 6.33 × 10−6 |
| BIEC2-600796 | Icelandic horse | 22 | 46,231,600 | L_Culo4 | A | B | 0.33 | 0.4650.114 | 6.49 × 10−5 |
| BIEC2_626250 | Shetland pony | 23 | 48,348,163 | L_Culo4 | A | B | 0.28 | 0.3270.089 | 2.47 × 10−4 |
| BIEC2_626256 | Shetland pony | 23 | 48,348,439 | L_Culo7 | B | A | 0.25 | 0.4230.114 | 2.58 × 10−4 |
| BIEC2_658221 | Shetland pony | 23 | 48,411,716 | L_Culo6 | A | B | 0.32 | −0.3400.092 | 2.33 × 10−4 |
| BIEC2_658438 | Shetland pony | 23 | 49,092,678 | L_Culo6 | A | B | 0.31 | −0.3530.093 | 1.38 × 10−4 |
| BIEC2_706297 | Shetland pony | 25 | 35,644,297 | L_Culo1b | B | A | 0.13 | −0.4040.109 | 2.01 × 10−4 |
| AX-103278700 | Belgian Warmblood horse | 26 | 7,825,717 | L_Culo1a | T | C | 0.22 | −1.0010.233 | 1.78 × 10−5 |
| AX-103417398 | Belgian Warmblood horse | 26 | 9,338,672 | L_Culo2a | A | G | 0.09 | −1.8310.383 | 1.72 × 10−6 |
| BIEC2_686588 | Icelandic horse | 26 | 16,094,903 | L_Culo4 | B | A | 0.20 | 0.5340.136 | 1.15 × 10−4 |
| BIEC2-692781 | Icelandic horse | 26 | 30,306,210 | L_Culo3 | A | B | 0.51 | −0.6130.148 | 3.44 × 10−5 |
| AX-104737542 | Belgian Warmblood horse | 27 | 23,911,229 | L_Culn5 | T | C | 0.46 | 0.8350.181 | 5.88 × 10−6 |
| AX-103948838 | Belgian Warmblood horse | 29 | 12,952,739 | L_Culn10 | T | C | 0.06 | 1.7280.400 | 1.53 × 10−5 |
Appendix D

Appendix E

Appendix F
| Study | Breed | Chr 1 | Pos (Mb) 2 |
|---|---|---|---|
| Current study | Icelandic horse | 1 | 62 |
| Current study | Icelandic horse | 1 | 64 |
| Current study | Shetland pony | 1 | 81 |
| Current study | Belgian Warmblood horse | 1 | 100 |
| Velie and colleagues [18] | Exmoor pony | 1 | 100 |
| Current study | Icelandic horse | 1 | 107 |
| Current study | Icelandic horse | 1 | 109 |
| Current study | Icelandic horse | 1 | 111 |
| Schurink and colleagues [15] | Shetland pony | 1 | 116 |
| Current study | Belgian Warmblood horse | 1 | 155 |
| Current study | Icelandic horse | 2 | 16 |
| Current study | Icelandic horse | 2 | 52 |
| Current study | Shetland pony | 2 | 53 |
| Current study | Icelandic horse | 2 | 55 |
| Current study | Belgian Warmblood horse | 2 | 57 |
| Current study | Icelandic horse | 2 | 63 |
| Current study | Shetland pony | 2 | 64 |
| Current study | Shetland pony | 2 | 65 |
| Current study | Shetland pony | 2 | 78 |
| Current study | Shetland pony | 2 | 89 |
| Current study | Shetland pony | 2 | 91 |
| Schurink and colleagues [16] | Friesian horse | 2 | 105 |
| Current study | Belgian Warmblood horse | 2 | 107 |
| Velie and colleagues [18] | Exmoor pony | 2 | 116 |
| Schurink and colleagues [14] | Shetland pony | 3 | 8 |
| Schurink and colleagues [15] | Shetland pony | 3 | 8 |
| Current study | Belgian Warmblood horse | 3 | 15 |
| Current study | Icelandic horse | 3 | 17 |
| Current study | Shetland pony | 3 | 17 |
| Schurink and colleagues [14] | Shetland pony | 3 | 17 |
| Schurink and colleagues [14] | Shetland pony | 3 | 50–51 |
| Current study | Shetland pony | 3 | 54 |
| Schurink and colleagues [15] | Shetland pony | 3 | 64 |
| Current study | Shetland pony | 3 | 71 |
| Current study | Icelandic horse | 3 | 78 |
| Shrestha and colleagues [17] | Icelandic horse | 3 | 87 |
| Velie and colleagues [18] | Exmoor pony | 4 | 1 |
| Current study | Shetland pony | 4 | 4 |
| Schurink and colleagues [14] | Icelandic horse | 4 | 43 |
| Current study | Belgian Warmblood horse | 4 | 44 |
| Current study | Icelandic horse | 5 | 25–26 |
| Schurink and colleagues [14] | Icelandic horse | 5 | 26 |
| Shrestha and colleagues [17] | Icelandic horse | 5 | 71 |
| Current study | Belgian Warmblood horse | 5 | 74 |
| Current study | Belgian Warmblood horse | 5 | 78 |
| Current study | Shetland pony | 5 | 98 |
| Current study | Shetland pony | 6 | 1 |
| Schurink and colleagues [14] | Icelandic horse | 6 | 6 |
| Current study | Icelandic horse | 6 | 6 |
| Current study | Shetland pony | 6 | 13 |
| Current study | Shetland pony | 6 | 15 |
| Current study | Belgian Warmblood horse | 6 | 44–46 |
| Schurink and colleagues [15] | Shetland pony | 6 | 47 |
| Shrestha and colleagues [17] | Icelandic horse | 6 | 48 |
| Velie and colleagues [18] | Exmoor pony | 6 | 55 |
| Current study | Belgian Warmblood horse | 6 | 59 |
| Current study | Icelandic horse | 6 | 73 |
| Current study | Belgian Warmblood horse | 6 | 79 |
| Schurink and colleagues [15] | Shetland pony | 7 | 13 |
| Current study | Icelandic horse | 7 | 19 |
| Schurink and colleagues [14] | Icelandic horse | 7 | 55 |
| Current study | Icelandic horse | 7 | 56 |
| Shrestha and colleagues [17] | Icelandic horse | 7 | 57 |
| Schurink and colleagues [15] | Shetland pony | 7 | 61 |
| Current study | Belgian Warmblood horse | 7 | 82 |
| Current study | Shetland pony | 7 | 85 |
| Schurink and colleagues [14] | Shetland pony | 7 | 85 |
| Current study | Icelandic horse | 8 | 27 |
| Velie and colleagues [18] | Exmoor pony | 8 | 62 |
| Schurink and colleagues [14] | Shetland pony | 8 | 63 |
| Current study | Belgian Warmblood horse | 8 | 64 |
| Schurink and colleagues [15] | Shetland pony | 8 | 70 |
| Velie and colleagues [18] | Exmoor pony | 8 | 70–71 |
| Velie and colleagues [18] | Exmoor pony | 8 | 78–79 |
| Current study | Icelandic horse | 8 | 79 |
| Current study | Shetland pony | 8 | 82 |
| Current study | Shetland pony | 8 | 92 |
| Velie and colleagues [18] | Exmoor pony | 9 | 26 |
| Current study | Shetland pony | 9 | 27 |
| Schurink and colleagues [16] | Friesian horse | 10 | 12–13 |
| Shrestha and colleagues [17] | Icelandic horse | 10 | 19 |
| Current study | Belgian Warmblood horse | 10 | 19 |
| Current study | Icelandic horse | 10 | 21 |
| Shrestha and colleagues [17] | Icelandic horse | 10 | 30 |
| Shrestha and colleagues [17] | Icelandic horse | 10 | 35 |
| Shrestha and colleagues [17] | Icelandic horse | 10 | 41 |
| Shrestha and colleagues [17] | Icelandic horse | 10 | 49 |
| Current study | Shetland pony | 10 | 51 |
| Current study | Shetland pony | 10 | 64 |
| Current study | Icelandic horse | 10 | 76 |
| Schurink and colleagues [14] | Shetland pony | 11 | 32 |
| Schurink and colleagues [15] | Shetland pony | 11 | 32–33 |
| Current study | Icelandic horse | 11 | 40 |
| Schurink and colleagues [14] | Icelandic horse | 11 | 40 |
| Schurink and colleagues [16] | Friesian horse | 11 | 41 |
| Current study | Belgian Warmblood horse | 11 | 43 |
| Current study | Belgian Warmblood horse | 12 | 7 |
| Schurink and colleagues [16] | Friesian horse | 12 | 19 |
| Current study | Belgian Warmblood horse | 12 | 19 |
| Current study | Belgian Warmblood horse | 13 | 2 |
| Current study | Belgian Warmblood horse | 13 | 11 |
| Current study | Icelandic horse | 13 | 17 |
| Current study | Shetland pony | 14 | 4 |
| Current study | Shetland pony | 14 | 13 |
| Current study | Shetland pony | 14 | 54 |
| Current study | Belgian Warmblood horse | 14 | 55 |
| Schurink and colleagues [16] | Friesian horse | 14 | 64 |
| Current study | Icelandic horse | 14 | 67–68 |
| Current study | Icelandic horse | 15 | 14 |
| Schurink and colleagues [14] | Icelandic horse | 15 | 19–20 |
| Current study | Icelandic horse | 15 | 21 |
| Current study | Belgian Warmblood horse | 15 | 31 |
| Schurink and colleagues [14] | Icelandic horse | 15 | 32–33 |
| Current study | Belgian Warmblood horse | 15 | 54 |
| Velie and colleagues [18] | Exmoor pony | 16 | 58–59 |
| Current study | Icelandic horse | 16 | 62 |
| Velie and colleagues [18] | Exmoor pony | 16 | 62–63 |
| Velie and colleagues [18] | Exmoor pony | 16 | 65 |
| Current study | Shetland pony | 17 | 19 |
| Current study | Shetland pony | 17 | 45 |
| Current study | Belgian Warmblood horse | 17 | 74–75 |
| Schurink and colleagues [14] | Shetland pony | 17 | 75–76 |
| Shrestha and colleagues [17] | Icelandic horse | 17 | 77 |
| Current study | Belgian Warmblood horse | 18 | 26 |
| Shrestha and colleagues [17] | Icelandic horse | 18 | 26 |
| Schurink and colleagues [14] | Icelandic horse | 18 | 32 |
| Current study | Belgian Warmblood horse | 18 | 58 |
| Current study | Icelandic horse | 19 | 39 |
| Schurink and colleagues [16] | Friesian horse | 20 | 19 |
| Schurink and colleagues [16] | Friesian horse | 20 | 24 |
| Schurink and colleagues [16] | Friesian horse | 20 | 26 |
| Current study | Shetland pony | 20 | 26 |
| Velie and colleagues [18] | Exmoor pony | 20 | 29 |
| Schurink and colleagues [14] | Icelandic horse | 20 | 30 |
| Schurink and colleagues [16] | Friesian horse | 20 | 29–30 |
| Schurink and colleagues [16] | Friesian horse | 20 | 30–32 |
| Schurink and colleagues [16] | Friesian horse | 20 | 32 |
| Current study | Shetland pony | 20 | 32 |
| Velie and colleagues [18] | Exmoor pony | 20 | 33 |
| Current study | Shetland pony | 20 | 34–35 |
| Schurink and colleagues [14] | Shetland pony | 20 | 35 |
| Schurink and colleagues [14] | Shetland pony | 20 | 41 |
| Schurink and colleagues [15] | Shetland pony | 20 | 41 |
| Current study | Shetland pony | 20 | 42 |
| Velie and colleagues [18] | Exmoor pony | 20 | 49 |
| Current study | Shetland pony | 20 | 51 |
| Current study | Belgian Warmblood horse | 20 | 52 |
| Schurink and colleagues [16] | Friesian horse | 21 | 10 |
| Current study | Shetland pony | 21 | 30 |
| Current study | Icelandic horse | 21 | 31 |
| Current study | Belgian Warmblood horse | 22 | 1 |
| Current study | Icelandic horse | 22 | 4 |
| Current study | Belgian Warmblood horse | 22 | 41 |
| Current study | Icelandic horse | 22 | 46 |
| Schurink and colleagues [16] | Friesian horse | 23 | 32 |
| Current study | Shetland pony | 23 | 48–49 |
| Current study | Shetland pony | 25 | 35 |
| Schurink and colleagues [16] | Friesian horse | 26 | 1 |
| Shrestha and colleagues [17] | Icelandic horse | 26 | 3 |
| Current study | Belgian Warmblood horse | 26 | 7 |
| Current study | Belgian Warmblood horse | 26 | 9 |
| Velie and colleagues [18] | Exmoor pony | 26 | 14 |
| Current study | Icelandic horse | 26 | 16 |
| Current study | Icelandic horse | 26 | 30 |
| Schurink and colleagues [15] | Shetland pony | 27 | 13 |
| Schurink and colleagues [14] | Shetland pony | 27 | 13 |
| Current study | Belgian Warmblood horse | 27 | 23 |
| Current study | Belgian Warmblood horse | 29 | 12 |
References
- Fadok, V.A.; Greiner, E.C. Equine insect hypersensitivity: Skin test and biopsy results correlated with clinical data. Equine Vet. J. 1990, 22, 236–240. [Google Scholar] [CrossRef] [PubMed]
- Gortel, K. Equine parasitic hypersensitivity—A review. Equine Pract. 1998, 20, 14–16. [Google Scholar]
- Riek, R.F. Studies on allergic dermatitis (Queensland itch) for the horse the aetiology of the disease. Aust. J. Agric. Res. 1954, 5, 109–129. [Google Scholar] [CrossRef]
- Eriksson, S.; Grandinson, K.; Fikse, W.F.; Lindgren, L.; Mikko, S.; Broström, H.; Frey, R.; Sundquist, M.; Lindgren, G. Genetic analysis of insect bite hypersensitivity (summer eczema) in Icelandic horses. Animal 2008, 2, 360–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schurink, A.; Van Grevenhof, E.M.; Ducro, B.J.; Van Arendonk, J.A.M. Heritability and repeatability of insect bite hypersensitivity in Dutch Shetland breeding mares. J. Anim. Sci. 2009, 87, 484–490. [Google Scholar] [CrossRef] [PubMed]
- Peeters, L.M.; Janssens, S.; Brebels, M.; Buys, N. Genetic parameters and estimated breeding values of insect bite hypersensitivity in Belgian Warmblood horses. Vet. J. 2015, 206, 420–422. [Google Scholar] [CrossRef] [PubMed]
- Björnsdóttir, S.; Sigvaldadóttir, J.; Broström, H.; Langvad, B.; Sigurdsson, A. Summer eczema in exported Icelandic horses: Influence of environmental and genetic factors. Acta Vet. Scand. 2006, 48, 3. [Google Scholar] [CrossRef]
- Broström, H.; Larsson, A.; Troedsson, M. Allergic dermatitis (sweet itch) of Icelandic horses in Sweden: An epidemiological study. Equine Vet. J. 1987, 19, 229–236. [Google Scholar] [CrossRef]
- Sommer-Locher, B.; Endriss, V.; Fromm, E. Various circumstances regarding initial allergen exposure and their influence on development of insect bite hypersensitivity in horses. J. Equine Vet. Sci. 2012, 32, 158–163. [Google Scholar] [CrossRef]
- Halldórdsóttir, S.; Larsen, H.J. An epidemiological study of summer eczema in Icelandic horses in Norway. Equine Vet. J. 1991, 23, 296–299. [Google Scholar] [CrossRef]
- Lange, S. Untersuchung zur Vererbung des Sommerekzems Beim Islandpferd. Ph.D. Thesis, Tierärztliche Hochschule, Hannover, Germany, 2004. [Google Scholar]
- Schurink, A.; Ducro, B.J.; Heuven, H.C.M.; Van Arendonk, J.A.M. Genetic parameters of insect bite hypersensitivity in Dutch Friesian broodmares. J. Anim. Sci. 2011, 89, 1286–1293. [Google Scholar] [CrossRef]
- Andersson, L.S.; Swinburne, J.E.; Meadows, J.R.S.; Broström, H.; Eriksson, S.; Fikse, W.F.; Frey, R.; Sundquist, M.; Tseng, C.T.; Mikko, S.; et al. The same ELA class II risk factors confer equine insect bite hypersensitivity in two distinct populations. Immunogenetics 2012, 64, 201–208. [Google Scholar] [CrossRef]
- Schurink, A.; Wolc, A.; Ducro, B.J.; Frankena, K.; Garrick, D.J.; Dekkers, J.C.M.; Van Arendonk, J.A.M. Genome-wide association study of insect bite hypersensitivity in two horse populations in the Netherlands. Gen. Sel. Evol. 2012, 44, 31–43. [Google Scholar] [CrossRef]
- Schurink, A.; Ducro, B.J.; Bastiaansen, J.W.; Frankena, K.; Van Arendonk, J.A.M. Genome-wide association study of insect bite hypersensitivity in Dutch Shetland pony mares. Anim. Genet. 2013, 44, 44–52. [Google Scholar] [CrossRef]
- Schurink, A.; Da Silva, V.H.; Velie, B.D.; Dibbits, B.W.; Crooijmans, R.P.M.A.; François, L.; Janssens, S.; Stinckens, A.; Blott, S.; Buys, N.; et al. Copy number variations in Friesian horses and genetic risk factors for insect bite hypersensitivity. BMC Genet. 2018, 19, 49. [Google Scholar] [CrossRef]
- Shrestha, M.; Eriksson, S.; Schurink, A.; Andersson, L.S.; Sundquist, M.; Frey, R.; Broström, H.; Bergström, T.; Ducro, B.J.; Lindgren, G. Genome-wide association study of insect bite hypersensitivity in Swedish-born Icelandic horses. J. Hered. 2015, 106, 366–374. [Google Scholar] [CrossRef]
- Velie, B.D.; Shrestha, M.; François, L.; Schurink, A.; Tesfayonas, Y.G.; Stinckens, A.; Blott, S.; Ducro, B.J.; Mikko, S.; Thomas, R.; et al. Using an inbred horse breed in a high density genome-wide scan for genetic risk factors of insect bite hypersensitivity (IBH). PLoS ONE 2016, 11, e0152966. [Google Scholar] [CrossRef]
- Abbas, A.K.; Lichtman, A.H. Basic Immunology: Functions and Disorders of the Immune System, 2nd ed.; Saunders: Philadelphia, PA, USA, 2004. [Google Scholar]
- Ferroglio, E.; Pregel, P.; Accossato, A.; Taricco, I.; Bollo, E.; Rossi, L.; Trisciuoglio, A. Equine Culicoides hypersensitivity: Evaluation of a skin test and of humoral response. J. Vet. Med. Ser. A 2006, 53, 30–33. [Google Scholar] [CrossRef]
- Frey, R.; Bergvall, K.; Egenvall, A. Allergen-specific IgE in Icelandic horses with insect bite hypersensitivity and healthy controls, assessed by FcεR1α-based serology. Vet. Immunol. Immunopathol. 2008, 126, 102–109. [Google Scholar] [CrossRef]
- Hellberg, W.; Wilson, A.D.; Mellor, P.; Doherr, M.G.; Torsteinsdottir, S.; Zurbriggen, A.; Jungi, T.; Marti, E. Equine insect bite hypersensitivity: Immunoblot analysis of IgE and IgG subclass responses to Culicoides nubeculosus salivary gland extract. Vet. Immunol. Immunopathol. 2006, 113, 99–112. [Google Scholar] [CrossRef]
- Hellberg, W.; Mellor, P.S.; Torsteinsdóttir, S.; Marti, E. Insect bite hypersensitivity in the horse: Comparison of IgE-binding proteins in salivary gland extracts from Simulium vittatum and Culicoides nubeculosus. Vet. Immunol. Immunopathol. 2009, 132, 62–67. [Google Scholar] [CrossRef]
- Langner, K.F.A.; Darpel, K.E.; Drolet, B.S.; Fischer, A.; Hampel, S.; Heselhaus, J.E.; Mellor, P.S.; Mertens, P.P.S.; Leibold, W. Comparison of cellular and humoral immunoassays for the assessment of summer eczema in horses. Vet. Immunol. Immunopathol. 2008, 122, 126–137. [Google Scholar] [CrossRef]
- Schaffartzik, A.; Marti, E.; Torsteinsdottir, S.; Mellor, P.S.; Crameri, R.; Rhyner, C. Selective cloning, characterization, and production of the Culicoides nubeculosus salivary gland allergen repertoire associated with equine insect bite hypersensitivity. Vet. Immunol. Immunopathol. 2011, 139, 200–209. [Google Scholar] [CrossRef]
- Van Der Meide, N.M.A.; Meulenbroeks, C.; Van Altena, C.; Schurink, A.; Ducro, B.J.; Wagner, B.; Leibold, W.; Rohwer, J.; Jacobs, F.; Van Oldruitenborgh-Oosterbaan, M.M.S.; et al. Culicoides obsoletus extract relevant for diagnostics of insect bite hypersensitivity in horses. Vet. Immunol. Immunopathol. 2012, 149, 245–254. [Google Scholar] [CrossRef]
- Wilson, A.D.; Harwood, L.J.; Björnsdottir, S.; Marti, E.; Day, M.J. Detection of IgG and IgE serum antibodies to Culicoides salivary gland antigens in horses with insect dermal hypersensitivity (sweet itch). Equine Vet. J. 2001, 33, 707–713. [Google Scholar] [CrossRef]
- Wilson, A.D.; Heesom, K.J.; Mawby, W.J.; Mellor, P.S.; Russell, C.J. Identification of abundant proteins and potential allergens in Culicoides nubeculosus salivary glands. Vet. Immunol. Immunopathol. 2008, 122, 94–103. [Google Scholar] [CrossRef]
- Peeters, L.M.; Janssens, S.; Goddeeris, B.M.; De Keyser, K.; Wilson, A.D.; Kaufmann, C.; Schaffartzik, A.; Marti, E.; Buys, N. Evaluation of an IgE ELISA with Culicoides spp. extracts and recombinant salivary antigens for diagnosis of insect bite hypersensitivity in Warmblood horses. Vet. J. 2013, 198, 141–147. [Google Scholar] [CrossRef]
- Schurink, A.; Van Der Meide, N.M.A.; Savelkoul, H.F.J.; Ducro, B.J.; Tijhaar, E. Factors associated with Culicoides obsoletus complex spp.-specific IgE reactivity in Icelandic horses and Shetland ponies. Vet. J. 2014, 201, 395–400. [Google Scholar] [CrossRef]
- Van Der Meide, N.M.A.; Roders, N.; Van Oldruitenborgh-Oosterbaan, M.M.S.; Schaap, P.J.; Van Oers, M.M.; Leibold, W.; Savelkoul, H.F.J.; Tijhaar, E. Cloning and expression of candidate allergens from Culicoides obsoletus for diagnosis of insect bite hypersensitivity in horses. Vet. Immunol. Immunopathol. 2013, 153, 227–239. [Google Scholar] [CrossRef]
- Van Der Meide, N.M.A.; Savelkoul, H.F.J.; Meulenbroeks, C.; Ducro, B.J.; Tijhaar, E. Evaluation of a diagnostic ELISA for insect bite hypersensitivity in horses using recombinant Obsoletus complex allergens. Vet. J. 2014, 200, 31–37. [Google Scholar] [CrossRef]
- Peeters, L.M. Genetic and Immunological Analysis of Insect Bite Hypersensitivity in Belgian Warmblood Horses. Ph.D. Thesis, KU Leuven, Leuven, Belgium, 2013. [Google Scholar]
- Bush, W.S.; Moore, J.H. Chapter 11: Genome-wide association studies. PLoS Comput. Biol. 2012, 8, e1002822. [Google Scholar] [CrossRef]
- Karssen, L.C.; Van Duijn, C.M.; Aulchenko, Y.S. The GenABEL project for statistical genomics. F1000Research 2016, 5, 914. [Google Scholar] [CrossRef]
- Peeters, L.M.; Janssens, S.; Schaffartzik, A.; Marti, E.; Buys, N. Evaluation of IgE levels against Culicoides nubeculosus allergens in Belgian Warmblood horses. Commun. Agric. Appl. Biol. Sci. 2012, 77, 218–222. [Google Scholar]
- Schaffartzik, A.; Marti, E.; Crameri, R.; Rhyner, C. Cloning, production and characterization of antigen 5 like proteins from Simulium vittatum and Culicoides nubeculosus, the first cross-reactive allergen associated with equine insect bite hypersensitivity. Vet. Immunol. Immunopathol. 2010, 137, 76–83. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2014; Available online: https://www.r-project.org/ (accessed on 15 May 2019).
- Tian, C.; Gregersen, P.K.; Seldin, M.F. Accounting for ancestry: Population substructure and genome-wide association studies. Hum. Mol. Genet. 2008, 17, 143–150. [Google Scholar] [CrossRef]
- Price, A.L.; Zaitlen, N.A.; Reich, D.; Patterson, N. New approaches to population stratification in genome-wide association studies. Nat. Rev. Genet. 2010, 11, 459–463. [Google Scholar] [CrossRef] [Green Version]
- Edwards, B.J.; Haynes, C.; Levenstien, M.A.; Finch, S.J.; Gordon, D. Power and sample size calculations in the presence of phenotype errors for case/control genetic association studies. BMC Genet. 2005, 6, 18. [Google Scholar] [CrossRef]
- McCarthy, M.I.; Abecasis, G.R.; Cardon, L.R.; Goldstein, D.B.; Little, J.; Ioannidis, J.P.A.; Hirschhorn, J.N. Genome-wide association studies for complex traits: Consensus, uncertainty and challenges. Nat. Rev. Genet. 2008, 9, 356–369. [Google Scholar] [CrossRef]
- Pearson, T.A.; Manolio, T.A. How to interpret a genome-wide association study. JAMA 2008, 299, 1335–1344. [Google Scholar] [CrossRef]
- Galvan, A.; Ioannidis, J.P.; Dragani, T.A. Beyond genome-wide association studies: Genetic heterogeneity and individual predisposition to cancer. Trends Genet. 2010, 26, 132–141. [Google Scholar] [CrossRef]
- Schaffartzik, A.; Hamza, E.; Janda, J.; Crameri, R.; Marti, E.; Rhyner, C. Equine insect bite hypersensitivity: What do we know? Vet. Immunol. Immonupathol. 2012, 147, 113–126. [Google Scholar] [CrossRef]
- Baselgia, S.; Doherr, M.G.; Mellor, P.; Torsteinsdottir, S.; Jermann, T.; Zurbriggen, A.; Jungi, T.; Marti, E. Evaluation of an in vitro sulphidoleukotriene release test for diagnosis of insect bite hypersensitivity in horses. Equine Vet. J. 2006, 38, 40–46. [Google Scholar] [CrossRef]
- Wagner, B.; Childs, B.A.; Erb, H.N. A histamine release assay to identify sensitization to Culicoides allergens in horses with skin hypersensitivity. Vet. Immunol. Immunopathol. 2008, 126, 302–308. [Google Scholar] [CrossRef]
- Van Oldruitenborgh-Oosterbaan, M.M.S.; Van Poppel, M.; De Raat, I.J.; Van Den Boom, R.; Savelkoul, H.F.J. Intradermal testing of horses with and without insect bite hypersensitivity in the Netherlands using an extract of native Culicoides species. Vet. Dermatol. 2009, 20, 607–614. [Google Scholar] [CrossRef]
- Laske, N.; Niggemann, B. Does the severity of atopic dermatitis correlate with serum IgE levels? Pediatr. Allergy Immunol. 2004, 15, 86–88. [Google Scholar] [CrossRef]
- Jia, P.; Wang, L.; Meltzer, H.Y.; Zhao, Z. Pathway-based analysis of GWAS datasets: Effective but caution required. Int. J. Neuropsychopharmacol. 2011, 14, 567–572. [Google Scholar] [CrossRef]
- Wang, K.; Li, M.; Bucan, M. Pathway-based approaches for analysis of genome-wide association studies. Am. J. Hum. Genetic. 2007, 81, 1278–1283. [Google Scholar] [CrossRef]
- Purcell, S.M.; Wray, N.R.; Stone, J.L.; Visscher, P.M.; O’Donovan, M.C.; Sullivan, P.F.; Sklar, P.; International Schizophrenia Consortium. Common polygenic variation contributes to risk of schizophrenia and bipolar disorder. Nature 2009, 460, 748–752. [Google Scholar] [CrossRef]
- Mooney, M.A.; Nigg, J.T.; McWeeney, S.K.; Wilmot, B. Functional and genomic context in pathway analysis of GWAS data. Trends Genet. 2014, 30, 390–400. [Google Scholar] [CrossRef] [Green Version]
- De Roos, A.P.; Hayes, B.J.; Spelman, R.J.; Goddard, M.E. Linkage disequilibrium and persistence of phase in Holstein-Friesian, Jersey and Angus cattle. Genetics 2008, 179, 1503–1512. [Google Scholar] [CrossRef]
- Schurink, A.; Shrestha, M.; Eriksson, E.; Bosse, M.; Bovenhuis, H.; Back, W.; Johansson, A.M.; Ducro, B.J. The genomic makeup of nine horse populations sampled in the Netherlands. Genes 2019, 10, 480. [Google Scholar] [CrossRef]
- Anderson, G.S.; Belton, P.; Kleider, N. Hypersensitivity of horses in British Columbia to extracts of native and exotic species of Culicoides (Diptera: Ceratopogonidae). J. Med. Entomol. 1993, 30, 657–663. [Google Scholar] [CrossRef]
- Van Der Rijt, R.; Van Den Boom, R.; Jongema, Y.; Van Oldruitenborgh-Oosterbaan, M.M.S. Culicoides species attracted to horses with and without insect hypersensitivity. Vet. J. 2008, 178, 91–97. [Google Scholar] [CrossRef]
- Singleton, A.B.; Hardy, J.; Traynor, B.J.; Houlden, H. Towards a complete resolution of the genetic architecture of disease. Trends Genet. 2010, 26, 438–442. [Google Scholar] [CrossRef] [Green Version]
- McCue, M.E.; Bannasch, D.L.; Petersen, J.L.; Gurr, J.; Bailey, E.; Binns, M.; Distl, O.; Guérin, G.; Hasegawa, T.; Hill, E.W.; et al. A high density SNP array for the domestic horse and extant Perissodactyla: Utility for association mapping, genetic diversity, and phylogeny studies. PLoS Genet. 2012, 8, e1002451. [Google Scholar] [CrossRef]
- Marian, A.J. Molecular genetic studies of complex phenotypes. Transl. Res. 2012, 159, 64–79. [Google Scholar] [CrossRef] [Green Version]


| Breed | |||
|---|---|---|---|
| Allergen | Belgian Warmblood Horse | Icelandic Horse | Shetland Pony |
| Total | 214 | 127 | 199 |
| Culo1a 1,2 | 193 | − | − |
| Culo1b 1,2 | − | 127 | 199 |
| Culo2a 1,2 | 165 | − | − |
| Culo2b 1,2 | − | 127 | 199 |
| Culo3 1 | − | 127 | 199 |
| Culo4 1 | − | 127 | 199 |
| Culo5 1 | − | 127 | 199 |
| Culo6 1 | − | 127 | 199 |
| Culo7 1 | − | 127 | 199 |
| Combi1 3 | − | 127 | 199 |
| Combi2 3 | − | 124 | 163 |
| Culn1 4 | 142 | − | − |
| Culn3 4 | 187 | − | − |
| Culn4 4 | 194 | − | − |
| Culn5 4 | 123 | − | − |
| Culn7 4 | 167 | − | − |
| Culn8 4 | 137 | − | − |
| Culn10 4 | 157 | − | − |
| WBE 5 | 182 | − | − |
| WBE 6 | − | 124 | 176 |
| nubTH 7 | 135 | − | − |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
François, L.; Hoskens, H.; Velie, B.D.; Stinckens, A.; Tinel, S.; Lamberigts, C.; Peeters, L.; Savelkoul, H.F.J.; Tijhaar, E.; Lindgren, G.; et al. Genomic Regions Associated with IgE Levels against Culicoides spp. Antigens in Three Horse Breeds. Genes 2019, 10, 597. https://doi.org/10.3390/genes10080597
François L, Hoskens H, Velie BD, Stinckens A, Tinel S, Lamberigts C, Peeters L, Savelkoul HFJ, Tijhaar E, Lindgren G, et al. Genomic Regions Associated with IgE Levels against Culicoides spp. Antigens in Three Horse Breeds. Genes. 2019; 10(8):597. https://doi.org/10.3390/genes10080597
Chicago/Turabian StyleFrançois, Liesbeth, Hanne Hoskens, Brandon D. Velie, Anneleen Stinckens, Susanne Tinel, Chris Lamberigts, Liesbet Peeters, Huub F. J. Savelkoul, Edwin Tijhaar, Gabriella Lindgren, and et al. 2019. "Genomic Regions Associated with IgE Levels against Culicoides spp. Antigens in Three Horse Breeds" Genes 10, no. 8: 597. https://doi.org/10.3390/genes10080597