Tolerance and Excretion of the Mycotoxins Aflatoxin B1, Zearalenone, Deoxynivalenol, and Ochratoxin A by Alphitobius diaperinus and Hermetia illucens from Contaminated Substrates


Abstract
:1. Introduction
2. Results
2.1. Mycotoxin Concentration in Feed
2.2. Effect of Mycotoxins on Insect Development
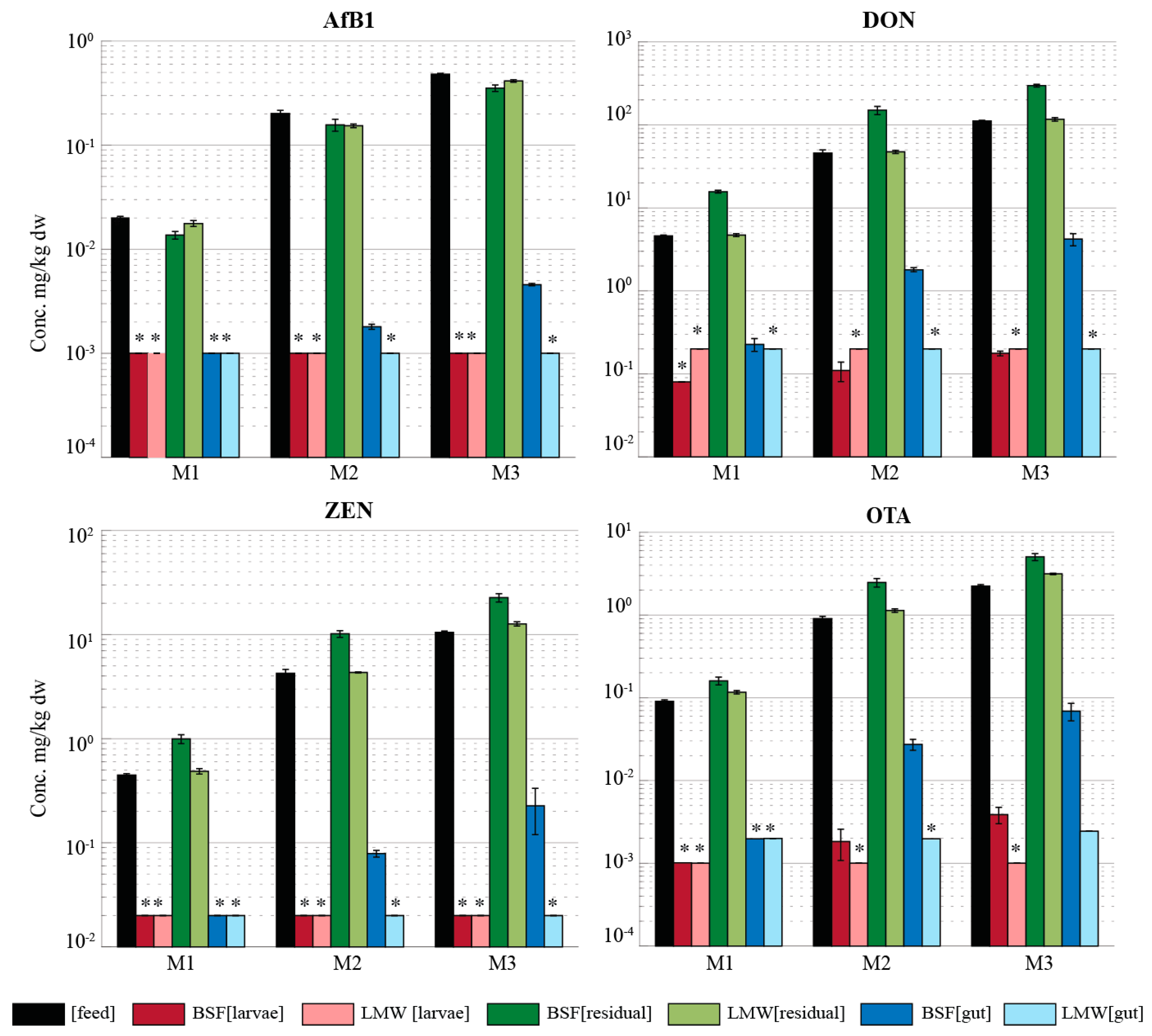
2.3. Mycotoxin Accumulation in Insects
2.4. Mycotoxin Metabolites
2.5. Mass Balance
3. Discussion
3.1. Insect Performance
3.2. Aflatoxin B1
3.3. Deoxynivalenol
3.4. Zearalenone and Ochratoxin A
3.5. Regulations
3.6. Further Research
4. Materials and Methods
4.1. Feed Preparation
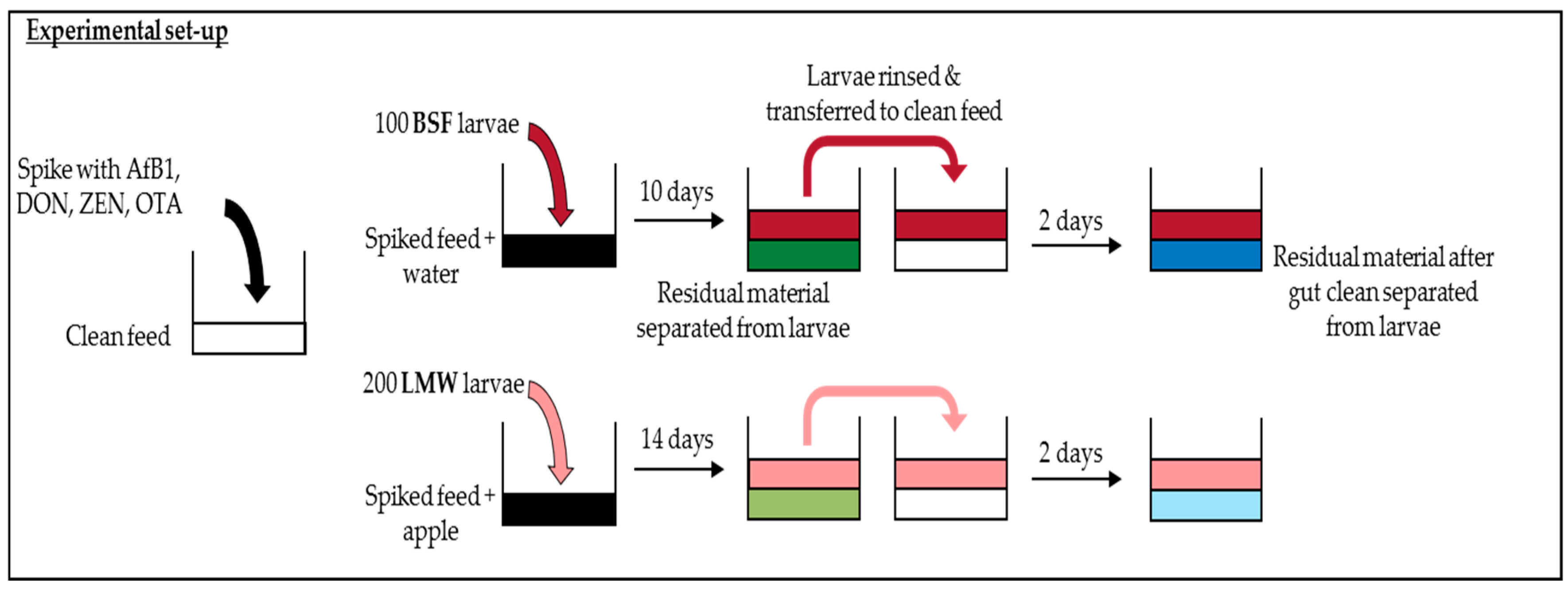
4.2. Experimental Set-Up
4.3. Sample Preparation and Mycotoxin Analyses
4.3.1. Standard and Chemicals
4.3.2. Extraction and Chemical Analyses
4.4. Data Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Van Raamsdonk, L.W.D.; Van der Fels-Klerx, H.J.; De Jong, J. New feed ingredients: The insect opportunity. Food Addit. Contam. 2017, 34, 1384–1397. [Google Scholar]
- European Commission. Commission Regulation (EC) No 1881/2006 of 19 December 2006 setting maximum levels for certain contaminants in foodstuffs. Off. J. Eur. Commun. 2006, 364/365, 1–20. [Google Scholar]
- European Commission. Commission Directive 2002/32/EC of the European Parliament and of the Council on undesirable substances in animal feed. Off. J. Eur. Commun. 2002, 140, 1–30. [Google Scholar]
- European Commission. Commission Recommendation of 17 August 2006 on the presence of deoxynivalenol, zearalenone, ochratoxin A, T-2 and HT-2 and fumonisins in products intended for animal feeding. Off. J. Eur. Commun. 2006, 229, 1–3. [Google Scholar]
- European Food Safety Authority. Risk profile related to production and consumption of insects as food and feed (EFSA-Q-2014-00578). EFSA J. 2015, 13, 4257. [Google Scholar]
- Van der Fels-Klerx, H.J.; Camenzuli, L.; van der Lee, M.K.; Oonincx, D.G.A.B. Uptake of cadmium, lead and arsenic by Tenebrio molitor and Hermetia illucens from contaminated substrates. PLoS ONE 2016, 11, e0166186. [Google Scholar] [CrossRef] [PubMed]
- Diener, S.; Zurbrügg, C.; Tockner, K. Bioaccumulation of heavy metals in the black soldier fly, Hermetia illucens and effects on its life cycle. J. Insects Food Feed 2015, 1, 261–270. [Google Scholar] [CrossRef]
- Houbraken, M.; Spranghes, T.; De Clercq, P.; Cooreman-Algoed, M.; Couchement, T.; De Clerct, G.; Verbeke, S.; Spanoghe, P. Pesticide contamination of Tenebrio molitor (Coleoptera: Tenebrionidea) for human consumption. Food Chem. 2016, 201, 264–269. [Google Scholar] [CrossRef] [PubMed]
- Van Broekhoven, S.; Gutierrez, J.M.; De Rijk, T.C.; De Nijs, W.C.M.; Van Loon, J.J.A. Degradation and excretion of the Fusarium toxin deoxynivalenol by an edible insect, the Yellow mealworm (Tenebrio molitor L.). World Mycotoxin J. 2017, 10, 163–169. [Google Scholar] [CrossRef]
- Bosch, G.; van der Fels-Klerx, H.J.; Rijk, T.C.D.; Oonincx, D.G.A.B. Aflatoxin B1 tolerance and accumulation in black soldier fly larvae (Hermetia illucens) and yellow mealworms (Tenebrio molitor). Toxins 2017, 9, 185. [Google Scholar] [CrossRef] [PubMed]
- Van Huis, A.; Van Itterbeeck, J.; Klunder, H.; Mertens, E.; Halloran, A.; Muir, G.; Vantomme, P. Edible Insects: Future Prospects for Food and Feed Security; Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 2013; p. 201. [Google Scholar]
- Purschke, B.; Scheibelberger, R.; Axmann, S.; Adler, A.; Jäger, H. Impact of substrate contamination with mycotoxins, heavy metals and pesticides on growth performance and composition of black soldier fly larvae (Hermetia illucens) for use in the feed and food value chain. Food Addit. Contam. 2017, 34, 1410–1420. [Google Scholar] [CrossRef] [PubMed]
- Tomberlin, J.K.; Sheppard, D.C.; Joyce, J.A. Selected life-history traits of black soldier flies (Diptera: Stratiomyidae) reared on three artificial diets. Ann. Entomol. Soc. Am. 2002, 95, 379–386. [Google Scholar] [CrossRef]
- Niu, G.; Siegel, J.; Schuler, M.A.; Berenbaum, M.R. Comparative toxicity of mycotoxins to navel orangeworm (Amyelois transitella) and corn earworm (Helicoverpa zea). J. Chem. Ecol. 2009, 35, 951–957. [Google Scholar] [CrossRef] [PubMed]
- Niu, G.; Johnson, R.M.; Berenbaum, M.R. Toxicity of mycotoxins to honeybees and its amelioration by propolis. Apidologie 2011, 42, 79–87. [Google Scholar] [CrossRef]
- Niu, G.; Wen, Z.; Rupasinghe, S.G.; Zeng, R.S.; Berenbaum, M.R.; Schuler, M.A. Aflatoxin B1 detoxification by CYP321A1 in Helicoverpa zea. Arch. Insect Biochem. Physiol. 2008, 69, 32–45. [Google Scholar] [CrossRef] [PubMed]
- Van Broekhoven, S.; Doan, Q.H.; van Huis, A.; van Loon, J.J. Exposure of tenebrionid beetle larvae to mycotoxin-contaminated diets and methods to reduce toxin levels. In Netherlands Entomological Society Meeting; FAO: Rome, Italy, 2014. [Google Scholar]
- European Food Safety Authority. Risks for animal health related to the presence of zearalenone and its modified forms in feed. EFSA J. 2017, 15, 4851. [Google Scholar] [CrossRef]
- Madrigal-Santillán, E.; Morales-González, J.A.; Vargas-Mendoza, N.; Reyes-Ramírez, P.; Cruz-Jaime, S.; Sumaya-Martínez, T.; Pérez-Pastén, R.; Madrigal-Bujaidar, E. Antigenotoxic Studies of different substances to reduce the DNA damage induced by aflatoxin B1 and ochratoxin A. Toxins 2010, 2, 738–757. [Google Scholar] [CrossRef] [PubMed]
- McCormick, S.P. Microbial detoxification of mycotoxins. J. Chem. Ecol. 2013, 39, 907–918. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mycotoxin Concentration (mg/kg ww) | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Level 1 | Level 2 | Level 3 | |||||||
| Mycotoxin | Intended | Analysed (Single) L1 | Analysed (Mix) M1 | Intended | Analysed (Single) L2 | Analysed (Mix) M2 | Intended | Analysed (Single) L3 | Analysed (Mix) M3 |
| AfB1 | 0.02 | 0.008 (16) | 0.018 (2.7) | 0.2 | 0.07 (13) | 0.18 (7.4) | 0.5 | 0.39 (12.0) | 0.43 (1.8) |
| DON | 5 | 3.9 (6.0) | 4.1 (2.4) | 50 | 38 (3.0) | 41 (9.0) | 125 | 112 (1.8) | 100 (1.3) |
| ZEN | 0.5 | 0.28 (4.9) | 0.4 (3.1) | 5 | 2.5 (2.5) | 3.8 (8.9) | 12.5 | 13 (5.1) | 9.4 (2.9) |
| OTA | 0.1 | 0.17 (4.0) | 0.08 (4.1) | 1 | 1.7 (7.0) | 0.8(6.5) | 2.5 | 1.3 (11.0) | 2 (4.3) |
| Aflatoxicol (Metabolite 1) | Aflatoxin M1 (Metabolite 2) | ||||||
|---|---|---|---|---|---|---|---|
| Larvae | Residual Material (Spiked Feed) | Residual Material (Gut Clean) | Larvae | Residual Material (Spiked Feed) | Residual Material (Gut Clean) | ||
| BSF | L1 | <0.001 b | <0.005 | <0.005 | <0.001 | <0.001 | <0.001 |
| L2 | <0.001 | <0.005 | <0.005 | <0.001 | <0.001 | <0.001 | |
| L3 | <0.001 | <0.005 | <0.005 | <0.001 | <0.001 | <0.001 | |
| M1 | <0.001 | <0.005 | <0.005 | <0.001 | <0.001 | <0.001 | |
| M2 | <0.001 | <0.005 | <0.005 | <0.001 | <0.001 | <0.001 | |
| M3 | <0.001 | 0.067 ± 0.002 | <0.005 | <0.001 | <0.001 | <0.001 | |
| LMW | L1 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 |
| L2 | <0.001 | <0.001 | <0.001 | <0.001 | 0.0020 ± 0.0002 | <0.001 | |
| L3 | <0.001 | 0.0015 ± 0.000 | <0.001 | <0.001 | 0.011 ± 0.0001 | <0.001 | |
| M1 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | |
| M2 | <0.001 | <0.001 | <0.001 | <0.001 | 0.0055 ± 0.001 | <0.001 | |
| M3 | <0.001 | 0.0013 ± 0.0002 | <0.001 | <0.001 | 0.011 ± 0.001 | <0.001 | |
| α-Zearalenol (Metabolite 1) | β-Zearalenol (Metabolite 2) | ||||||
| BSF | L1 | <0.005 | 0.64 ± 0.026 | <0.02 | <0.005 | 0.18 ± 0.010 | <0.01 |
| L2 | 0.005 ± 0.002 | 7.2 ± 0.31 | 0.14 ± 0.04 | <0.005 | 2.3 ± 0.12 | 0.042 ± 0.02 | |
| L3 | 0.025 ± 0.004 | 37.3 ± 8.1 | 0.60 ± 0.18 | 0.007 ± 0.001 | 11.2 ± 1.32 | 0.18 ± 0.06 | |
| M1 | <0.005 | 0.74 ± 0.11 | 0.022 ± 0.002 | <0.005 | 0.20 ± 0.026 | <0.02 | |
| M2 | 0.011 ± 0.004 | 12.7 ± 2.9 | 0.21 ± 0.012 | <0.005 | 3.6 ± 0.85 | 0.071 ± 0.051 | |
| M3 | 0.029 ± 0.005 | 28.3 ± 1.5 | 0.63 ± 0.078 | 0.0067 ± 0.001 | 8.6 ± 1.02 | 0.13 ± 0.032 | |
| LMW | L1 | <0.005 | 0.023 ± 0.002 | <0.02 | <0.005 | <0.01 | <0.01 |
| L2 | <0.005 | 0.203 ± 0.012 | <0.02 | <0.005 | 0.102 ± 0.007 | <0.01 | |
| L3 | <0.005 | 0.82 ± 0.071 | <0.02 | <0.005 | 0.32 ± 0.026 | <0.01 | |
| M1 | <0.005 | 0.030 ± 0.002 | <0.02 | <0.005 | 0.011 ± 0.001 | <0.01 | |
| M2 | <0.005 | 0.35 ± 0.040 | <0.02 | <0.005 | 0.14 ± 0.012 | <0.01 | |
| M3 | <0.005 | 0.74 ± 0.012 | <0.02 | <0.005 | 0.27 ± 0.015 | <0.01 | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Camenzuli, L.; Van Dam, R.; De Rijk, T.; Andriessen, R.; Van Schelt, J.; Van der Fels-Klerx, H.J. Tolerance and Excretion of the Mycotoxins Aflatoxin B1, Zearalenone, Deoxynivalenol, and Ochratoxin A by Alphitobius diaperinus and Hermetia illucens from Contaminated Substrates. Toxins 2018, 10, 91. https://doi.org/10.3390/toxins10020091
Camenzuli L, Van Dam R, De Rijk T, Andriessen R, Van Schelt J, Van der Fels-Klerx HJ. Tolerance and Excretion of the Mycotoxins Aflatoxin B1, Zearalenone, Deoxynivalenol, and Ochratoxin A by Alphitobius diaperinus and Hermetia illucens from Contaminated Substrates. Toxins. 2018; 10(2):91. https://doi.org/10.3390/toxins10020091
Chicago/Turabian StyleCamenzuli, Louise, Ruud Van Dam, Theo De Rijk, Rob Andriessen, Jeroen Van Schelt, and H. J. (Ine) Van der Fels-Klerx. 2018. "Tolerance and Excretion of the Mycotoxins Aflatoxin B1, Zearalenone, Deoxynivalenol, and Ochratoxin A by Alphitobius diaperinus and Hermetia illucens from Contaminated Substrates" Toxins 10, no. 2: 91. https://doi.org/10.3390/toxins10020091