1. Introduction
Fluorescent proteins (FPs) are nowadays indispensable in life sciences [
1,
2,
3,
4]. The discovery of FPs started in the early 1960s with studies on the identification of the glow of jellyfish from
Aequorea victoria by Osamu Shimomura [
5]. The protein emitting the green light was called green fluorescent protein (GFP) [
6] and its sequence was obtained in 1992 by Prasher [
7]. In the following years, a wide variety of GFP variants with different colors and improved brightness and stability were developed. However, there were no GFP variants with emission maxima above 527 nm [
4]. This limitation was overcome by cloning of GFP homologs from non-bioluminescent reef corals of the Anthozoa class [
8,
9,
10,
11]. From this class, a palette of FPs became available emitting at longer wavelengths. Consequently, the number of applications of FPs has exploded, which is mainly because they can be genetically introduced into cells, tissues or whole organisms. This allows using FPs for multicolor imaging and for studying protein interactions [
12,
13,
14]. Besides using FPs as fusion tags and biosensors, they have also been used as model proteins in encapsulation studies [
15,
16,
17,
18].
Encapsulation of proteins is of interest for food and biomedical applications, because it can protect and stabilize the encapsulated protein. Encapsulation of FPs allows the use of fluorescence techniques for the characterization of protein-containing micelles [
18].
Previously, we reported on the encapsulation of enhanced green fluorescent protein (EGFP) and its monomeric variant (mEGFP) in complex coacervate core micelles (C3Ms) with the cationic-neutral diblock copolymer poly(2-methyl-vinyl-pyridinium)
n-
b-poly(ethylene-oxide)
m (P2MVP
n-
b-PEO
m) as enveloping material [
18,
19]. The two GFP variants showed considerable differences in their spectral and structural properties upon encapsulation. Encapsulation into C3Ms promoted dimerization of EGFP but not of mEGFP, due to the difference in dissociation constant (
KD, 0.11 mM for EGFP and 74.0 mM for mEGFP [
20]). Dimerization of EGFP upon encapsulation in C3Ms results in a p
Ka shift of its chromophore, leading to specific changes in the spectral and structural properties of EGFP [
19].
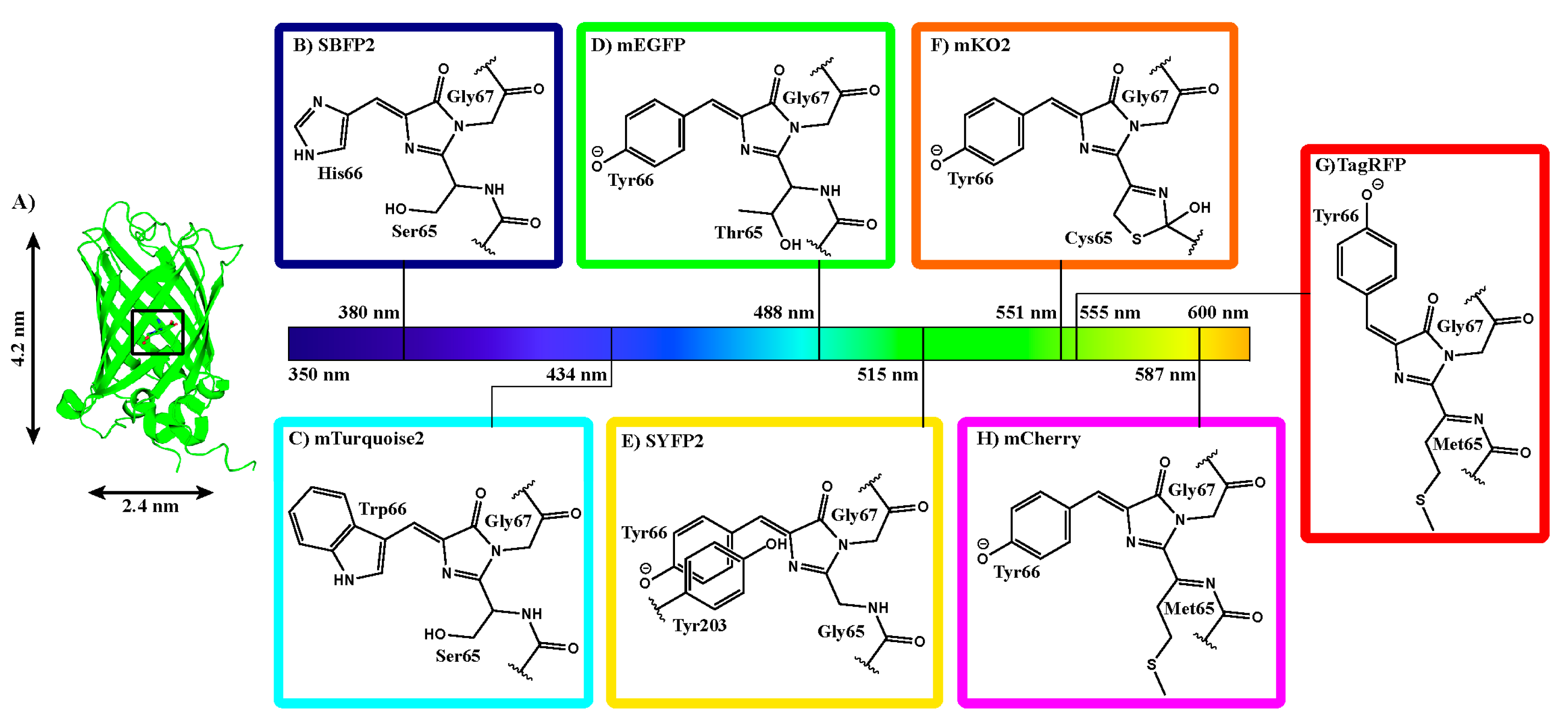
The aim of the present study is to determine whether structural and spectral changes are common upon encapsulation of members of the visible fluorescent protein family. Therefore, we encapsulated a variety of fluorescent proteins covering the whole visible spectrum (
Figure 1). We investigated seven differently colored FPs: four FPs derived from
Aequorea victoria GFP (
avFPs: strongly enhanced blue fluorescent protein 2 (SBFP2), mTurquoise2, mEGFP, and strongly enhanced yellow fluorescent protein 2 (SYFP2), from class Hydrozoa) and three FPs from class Anthozoa (
anFPs: monomeric Kusabira Orange 2 (mKO2), Tag red fluorescent protein (TagRFP), and mCherry). The seven FPs originate from four different ancestors: one from the Hydrozoa class and three from the Anthozoa class. Even though these proteins have a comparable fold and function, the amino acid sequences vary between the different species (see
Figure S1 and Table S1). The sequence identities among the
avFPs are about 97%, whereas the
anFPs show about 25–30% similarity to the
avFPs. Amongst the
anFPs, TagRFP and mCherry show a higher sequence identity (57%) than mKO2 does with these two proteins (~48%). The seven FPs contain about 10% of strictly conserved residues (
Figure S1). These residues are mainly located at the end of beta-strands and in loops, except for the strictly conserved Gly67, Asp95, Arg96, and Glu222 amino acid residues, which are involved in the formation of the chromophore [
21].
Some general properties of FPs are critical for understanding their structural and spectral properties. All FPs have an approximate molar mass of 27 kDa, constituting a single polypeptide chain of about 230 amino acids. They all share a fold consisting of an 11-stranded β-barrel with a length of 4.2 nm and a diameter of 2.4 nm (
Figure 1A). The barrel is wrapped around a single distorted helix, which contains three amino acid residues that create the fluorophore. The fluorophore is formed via cyclization, dehydration, and oxidation of the amino acid residues located at positions 65–67 (mEGFP numbering,
Figure 1B–H). Depending on the pH of the solution, the chromophore can exist in differing protonation states, which influences the spectral properties of the FP. Furthermore, the fluorophore is comprised of a highly conjugated π-electron resonance system that together with its environment accounts for the spectroscopic and photophysical properties of an FP.
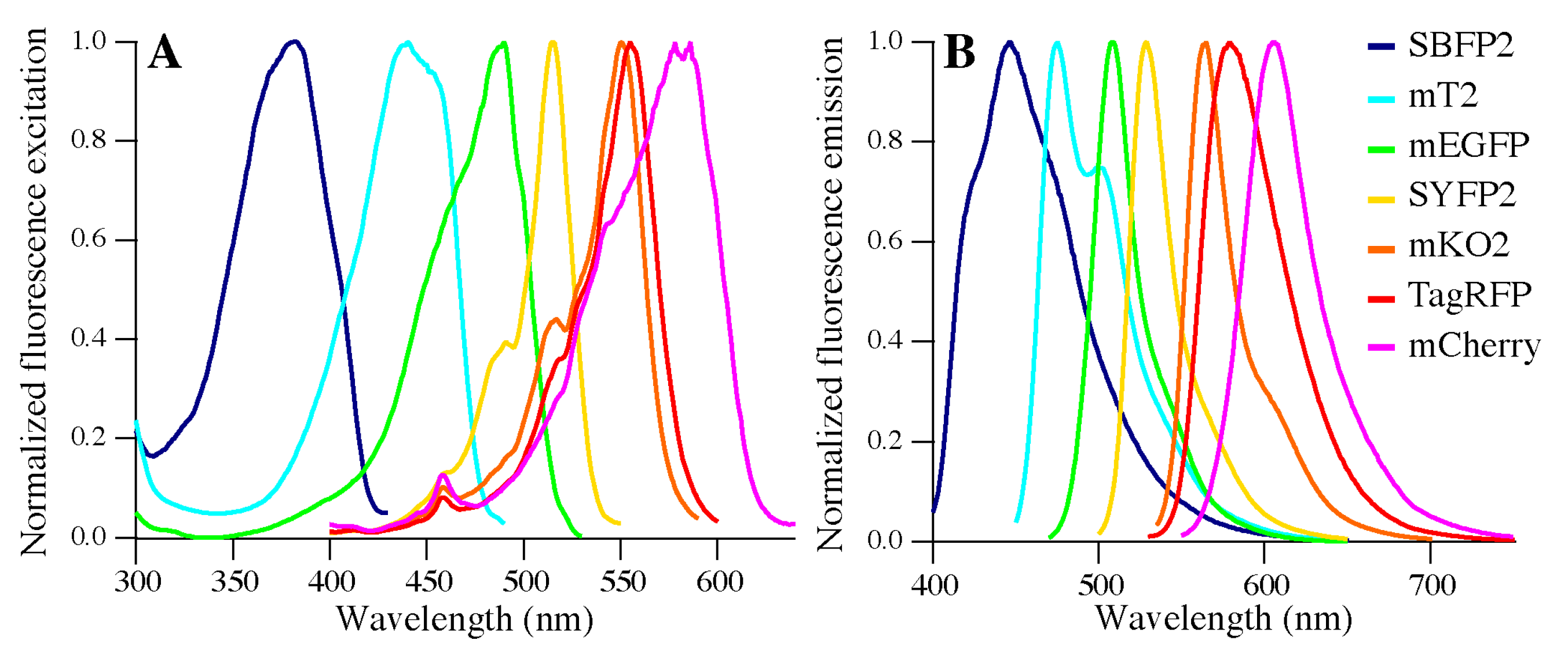
The spectroscopic features of the seven FPs will be presented according to their absorption maxima. We start with strongly enhanced blue fluorescent protein (SBFP2), which was obtained by the Y66H substitution in GFP (
Figure 1B) [
22]. Turquoise FPs are obtained by a Y66W substitution in GFP (
Figure 1C), which yields amongst others mTurquoise2 [
23]. mTurquoise2 is especially characterized by its long mono-exponential fluorescence lifetime (τ = 3.8 ns), which makes it very suitable as a Förster resonance energy transfer (FRET) donor in conjunction with a yellow fluorescent protein as the acceptor. Originally, FPs are not monomeric, but this can be achieved with the A206K substitution, which is used in mEGFP [
20]. The GFP variants with emission maxima at the longest wavelengths are yellow FPs, obtained by a T203Y substitution in GFP, and in this research we used SYFP2 (
Figure 1E) [
24]. This FP has a high extinction coefficient compared to other FPs (ε
SYFP2 = 101 000 M
−1·cm
−1, see
Table S2) making it a very suitable acceptor in FRET-pairs [
24].
The chromophore structures of FPs from Anthozoa species generally have more extended π-systems, enabling higher excitation and emission wavelengths. Such a type of fluorophore is found in mKO2, which evolved from a fluorescent protein of the mushroom coral
Fungia concinna, with a cysteine located at position 65 (mEGFP numbering,
Figure 1F) [
9,
25]. mKO2 is very useful in multicolor imaging applications as it can be combined with cyan, green, yellow, and red FPs. A protein with an almost similar excitation maximum as mKO2 is TagRFP (
Figure 1G), but this protein has been derived from the sea anemone
Entacmaea quadricolor [
10]. TagRFP has an even more extended π-system than mKO2, because it has a methionine located at position 65 (mEGFP numbering,
Figure 1G). Next to that, TagRFP is one of the few FPs bearing a
trans-isomerized chromophore. A protein that also contains a methionine at position 65 (mEGFP numbering) is mCherry, one of the “mFruit” FPs derived from
Discosoma species [
26,
27]. mCherry shows a high photostability and its chromophore is rapidly formed (
Figure 1H), which makes it very suitable as a FRET acceptor in combination with EGFP in fluorescence-lifetime imaging microscopy (FLIM) studies [
28].
In this research, we used the diblock copolymers P2MVP41-b-PEO205 and P2MVP128-b-PEO477 to form C3Ms in combination with the above-mentioned FPs. We characterized the C3Ms with dynamic light scattering (DLS) and fluorescence correlation spectroscopy (FCS), and explored the effects of packing on the FPs with circular dichroism (CD) and fluorescence spectral analysis. The experimental data, and in particular the observed encapsulation efficiencies, are discussed in relation to what is known about the structural features of the FPs.
3. Discussion
Previously, we found that the encapsulation of EGFP in C3Ms stimulates protein dimerization and changes the spectral properties of the EGFP chromophore [
19]. Because mEGFP mainly remains monomeric in the densely packed C3Ms, encapsulation of this protein hardly affects its spectral properties. In this work, we studied the encapsulation of four
avFPs and three
anFPs in C3Ms. All investigated FPs were successfully encapsulated using two diblock copolymers (P2MVP
41-
b-PEO
205 and P2MVP
128-
b-PEO
477) with
F+ values ranging between 0.60 and 0.80. For strong polyelectrolytes, stoichiometric C3M systems are formed at a
F+ value of 0.50 [
41]. Proteins, however, are weak polyelectrolytes and therefore their charge may change upon interaction with the diblock copolymer. Moreover, coacervation between polymer and protein does not necessarily arise from the overall charge of the protein, but rather from specific charge patches on the protein surface [
42]. Both effects can even lead to coacervation between similarly charged proteins and polyelectrolytes [
43,
44,
45,
46].
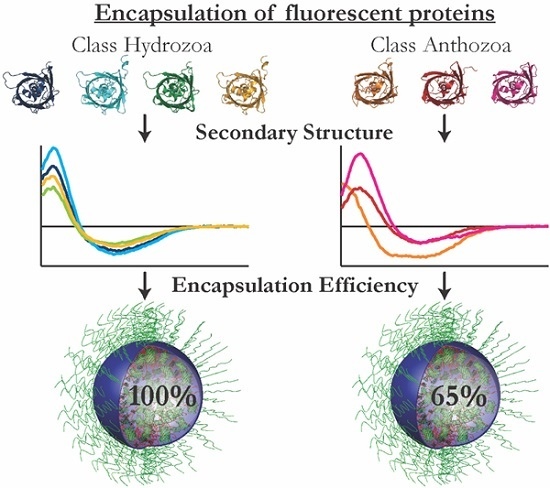
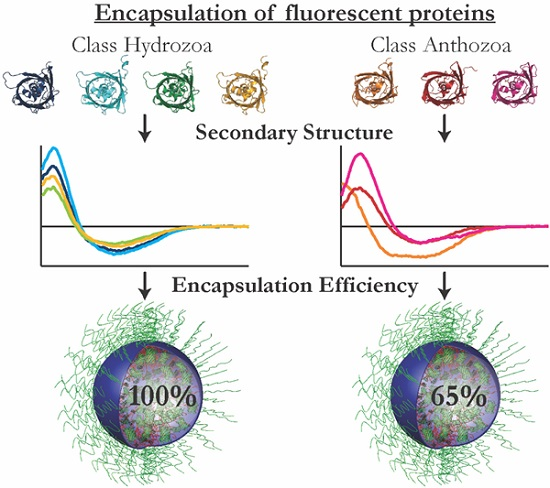
3.1. Encapsulation Efficiency
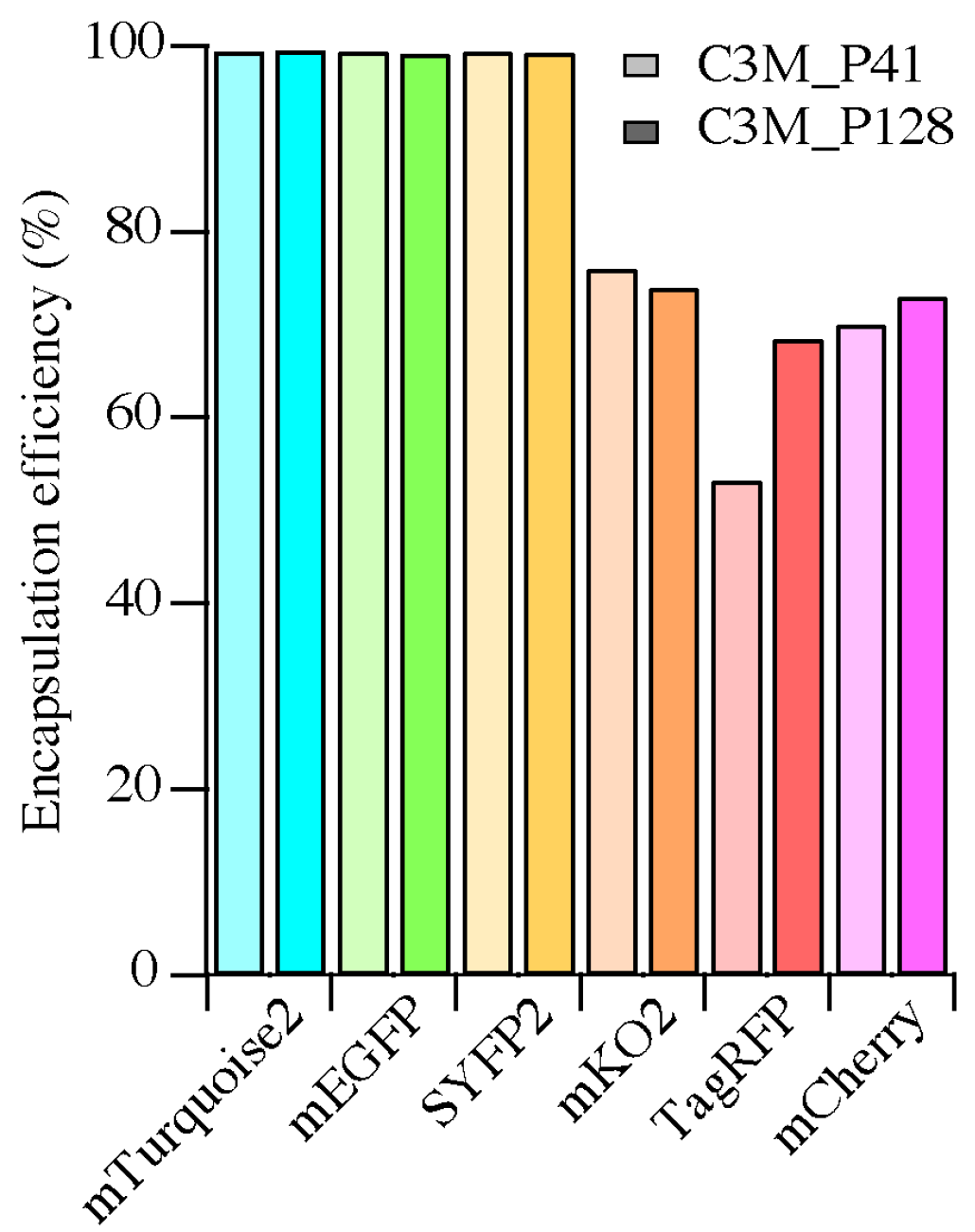
The encapsulation efficiencies of
avFPs (mEGFP, SBFP2, SYFP2 and mTurquoise2) were almost 100%, whereas those of
anFPs (mKO2, TagRFP and mCherry) varied between 50% and 75%. This implicates that the interactions between the
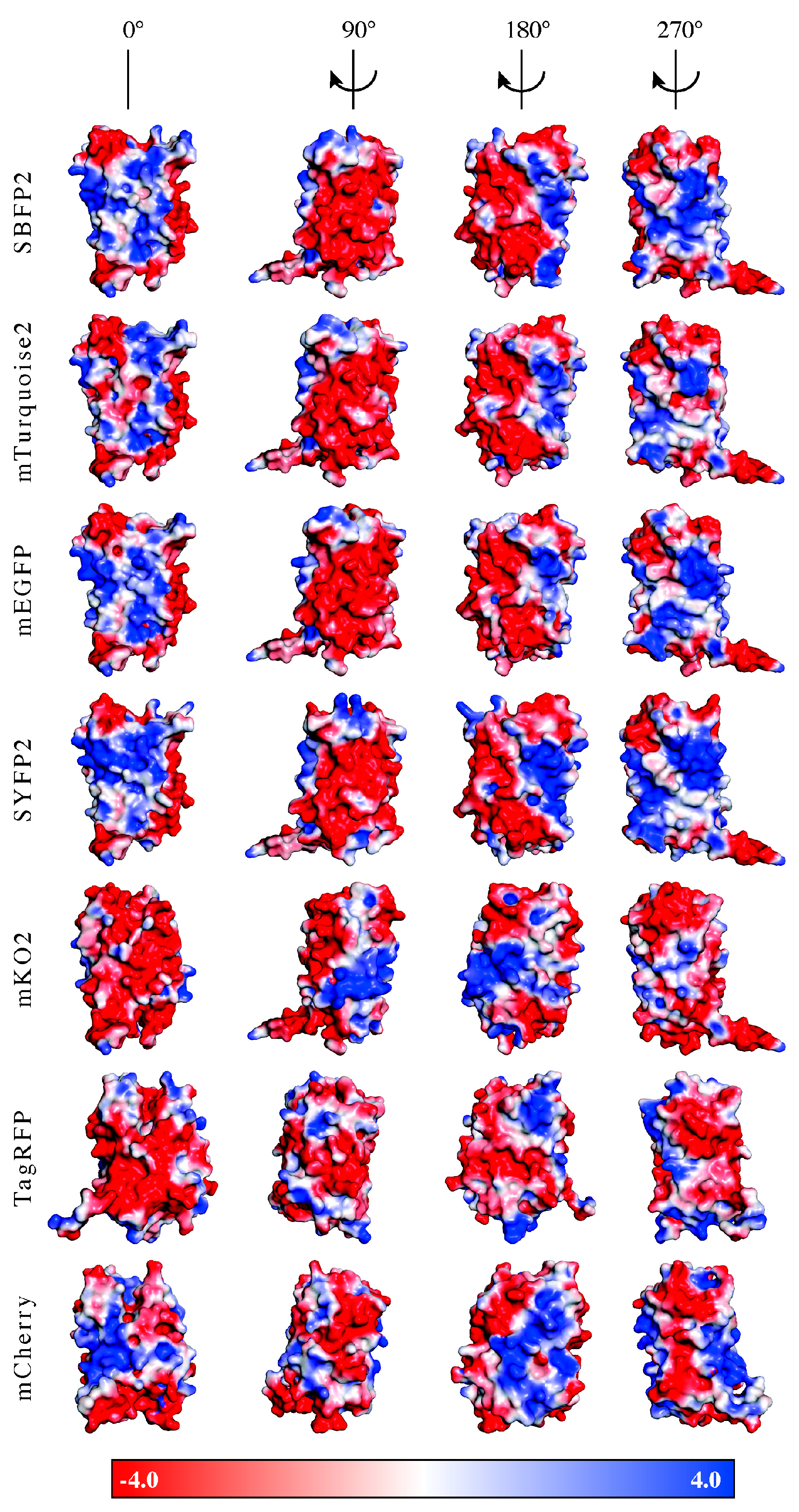
anFPs and the diblock copolymers to form C3Ms are less favorable. The formation of C3Ms requires an interaction between the FPs and the polymers, which can be dependent on the surface charge distribution and/or the shape of the protein. For the investigation of the presence of specific charge patches on the protein surface, we determined the surface potential distribution of the FPs on the acquired protein structures. For this, homology modeling was used to obtain the protein structures of SBFP2, SYFP2, and mKO2, next to the crystal structures of mTurquoise2, mEGFP, TagRFP, and mCherry. In
Figure 7, the surface potentials of the FPs are visualized at the pH value at which they were encapsulated. All
avFPs share a negative surface patch, as displayed on the side view at 90°, with an expansion to half of the molecule displayed in the side view at 180°. The amino acid residues with negative charge belonging to this patch are located on β-strands 1 and 2. The three
anFPs do not contain a similar negative patch displayed on the side view at 90°, as observed for
avFPs. Negative patches for mKO2 and TagRFP are mainly present in the side view at 0°. For TagRFP, the amino acid residues with negative charge are more distributed over the entire protein surface than for the other proteins. For mCherry, there is not a side entirely filled with negatively charged amino acid residues. It is key for the positively charged polyelectrolyte to bind to a local negative charge patch on the protein while minimizing the repulsive effect arising from the positively charged amino acid residues. Therefore, the interactions between the diblock copolymers and mKO2, TagRFP, and mCherry might not be optimal, thus affecting their encapsulation efficiencies.
3.2. Elliptical Symmetry of FP Barrels
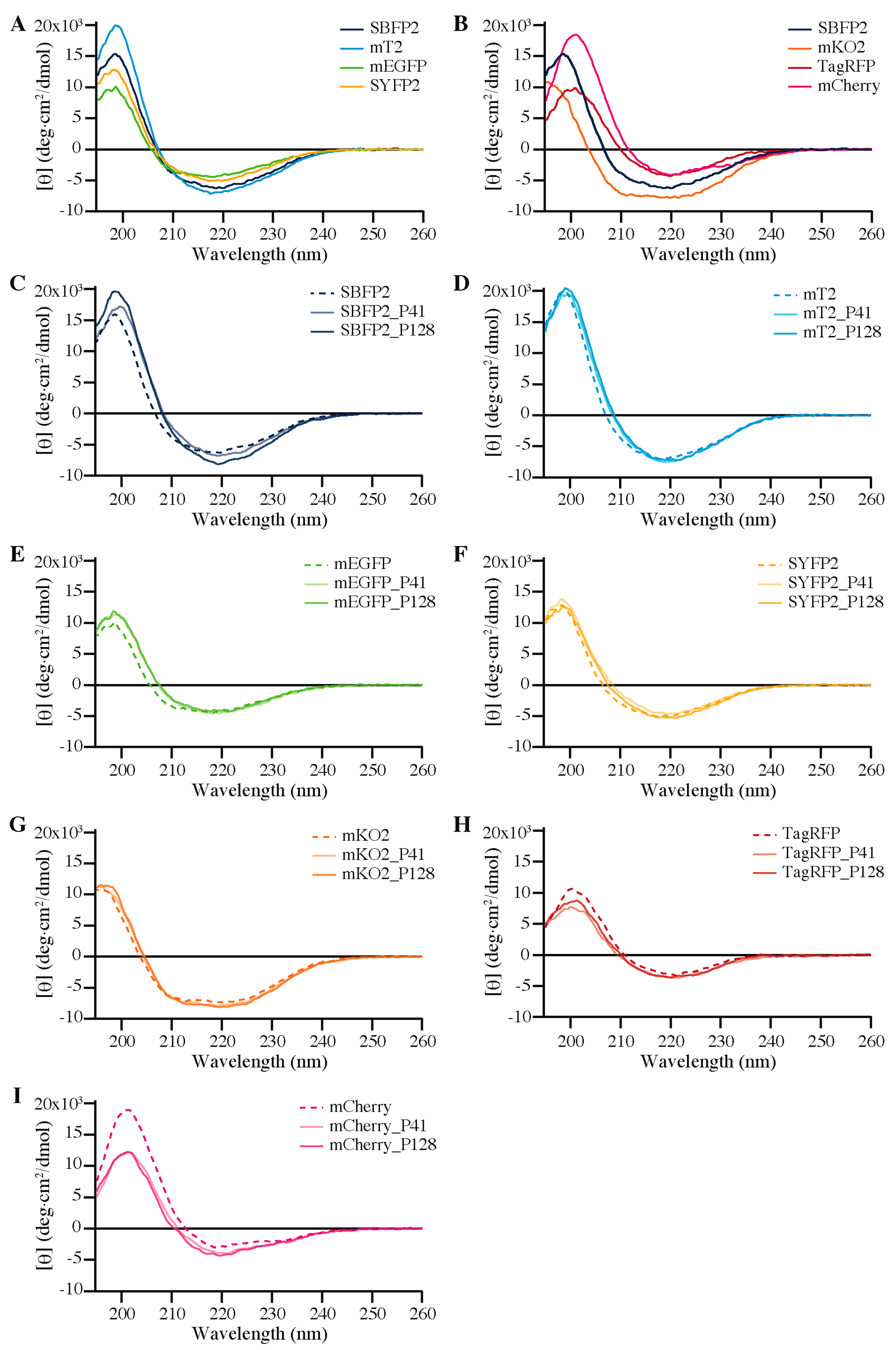
During this study, we uncovered clear differences in the far-UV CD spectra between
avFPs and
anFPs free in solution (
Figure 6B). It is well known that all FPs share a similar 11-stranded β-barrel fold (
Figure 1A). However, it is hardly reported in the literature that the elliptical symmetry between
avFPs and
anFPs is diverse [
47].
Figure 8 shows the ribbon structures of the studied FPs in three different orientations: the broad side, the narrow side, and the top. From the top views, it is clear that the barrels of the FPs are not completely round, but form elliptic cylinders. The
avFPs are rounder than the
anFPs, which is depicted by the difference in aspect ratio: ~0.85 for the
avFPs and ~0.74 for the
anFPs. Especially mKO2 is the most “squeezed” of the
anFPs. We hypothesize that these differences are the cause for the observed differences in the far-UV CD spectra. Micsonai et al. reported that CD spectra are influenced, among others, by degree of twist and distortion of the β-sheets [
48]. The variance in the elliptical symmetry is another apparent difference between
avFPs and
anFPs, and could also be influencing their encapsulation efficiencies.
3.3. Biophysical Properties of Encapsulated Proteins
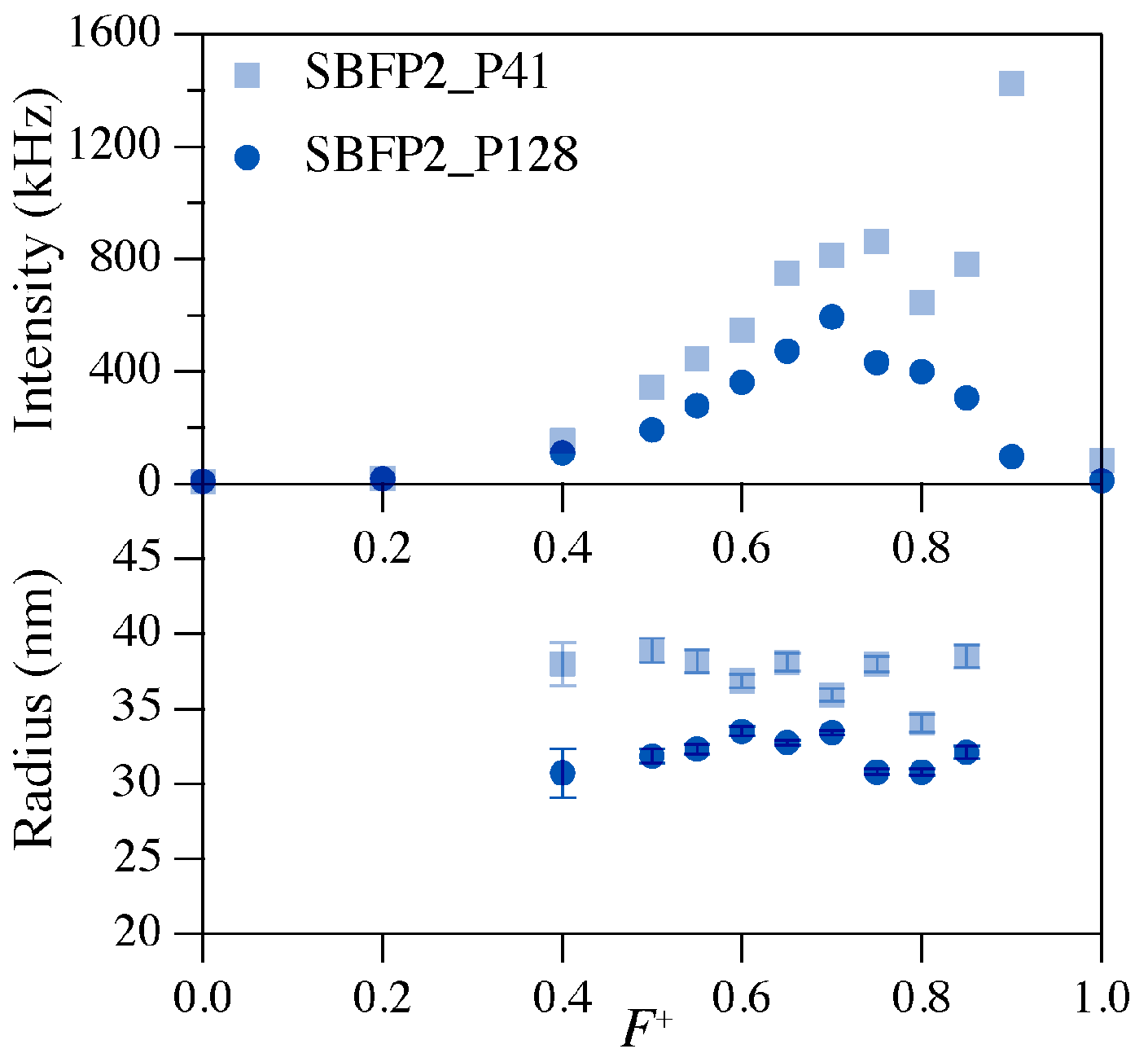
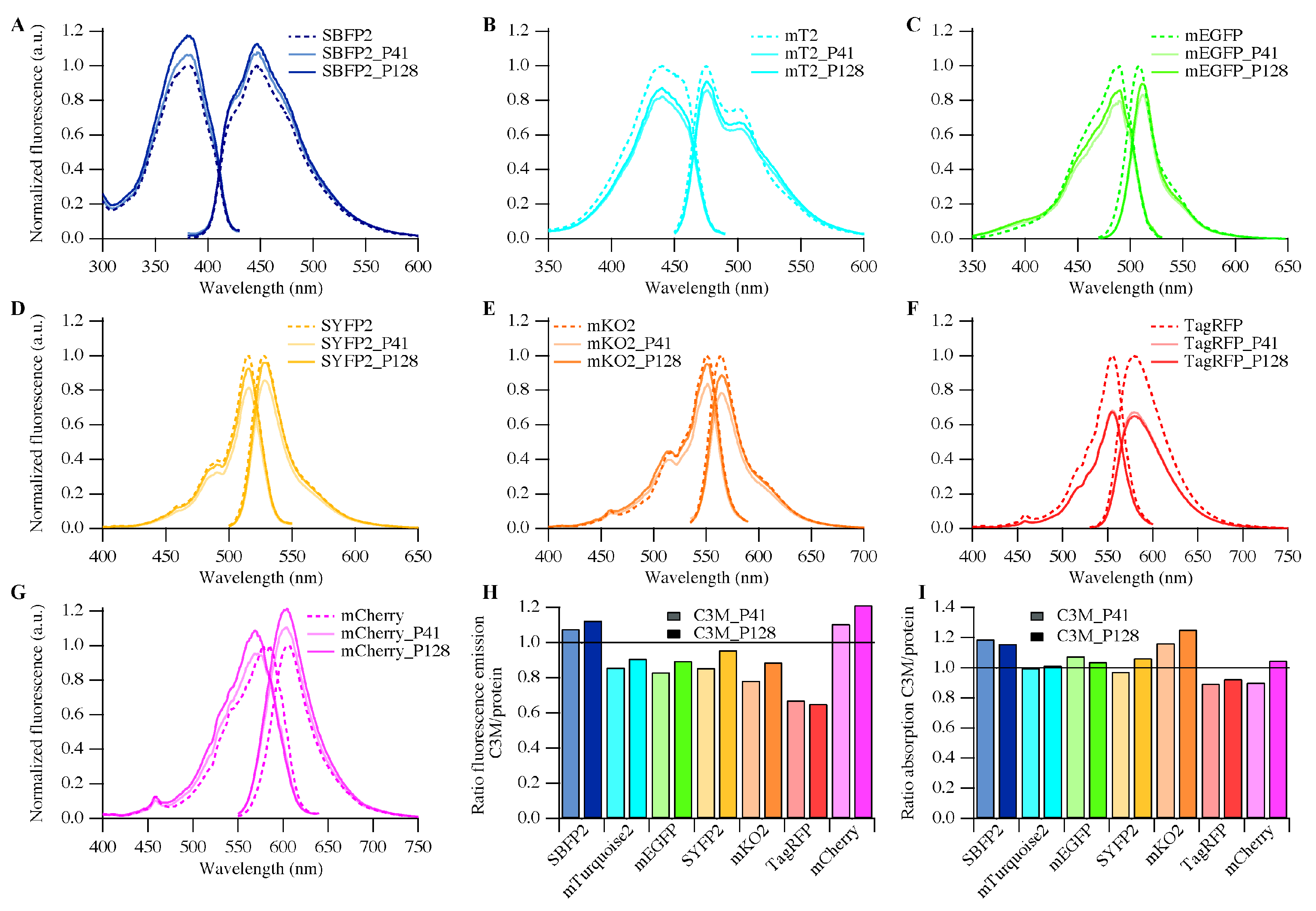
Encapsulation of the avFPs hardly influenced their secondary structural properties and only minor changes in absorption and emission characteristics were observed. All avFPs bear the A206K mutation, which favors their monomeric state. This supports that the minor spectral changes observed are caused by the electrostatic interactions between the polymers and the protein surfaces of these FPs.
All
anFPs are found as tetramers in their hosts [
9,
10,
26]. The
anFPs used here are all modified to enhance their tendency to remain monomeric. In literature, this tendency is expressed in terms of dissociation constants and “monomeric quality” (see
Table S2). Previously, we calculated the number of EGFP molecules present in a C3M to be around 400, yielding a local protein concentration of about 10 mM [
18,
19]. Since the FPs used in this study form C3Ms with PMCs (~0.65) and radii (~34 nm) similar to EGFP-C3Ms, it is a reasonable assumption that the protein concentration in the various FP-C3Ms is about the same. Hence, we expect that mCherry with a monomeric quality of 95% remains mostly monomeric upon encapsulation. However, mKO2 and TagRFP with monomeric qualities of 68% and 58%, respectively, and a dissociation constant of 0.038 mM for TagRFP, will likely form oligomers in the C3Ms (
Table S2). This oligomerization causes the large decrease in quantum yield of the encapsulated forms of mKO2 and TagRFP (
Table 2).
For encapsulated mCherry, the absorption spectrum changes according to a p
Ka shift of its chromophore (
Figure S4). For EGFP it was proposed that the p
Ka shift of its chromophore is caused by a reorientation of Glu222 due to the dimerization of EGPF in C3Ms [
19]. For free mCherry, the equivalent Glu215 is also linked to the pH-dependent spectral shifts (
Figure S5G) [
27]. If mCherry, however, remains monomeric in the C3Ms, the reorientation of Glu215 can only occur due to the interaction between protein and polymer.
3.4. Future Research
We show that encapsulation of structurally similar FPs in C3Ms is dependent on the origin of the FPs and can give rise to different encapsulation efficiencies. Moreover, the spectral and structural perturbations observed are dependent on the kind of FP and the type of polymer used. In future research, we plan to investigate the stability and dynamics of encapsulated FPs. This can be accomplished by mixing two appropriate FPs using FRET as a readout. Some requirements should be considered choosing an optimal FP FRET-pair: First, use fluorescent proteins with similar encapsulation efficiencies. Second, use FPs that show minor changes in their absorption and fluorescence properties upon encapsulation into the C3Ms. Third, use the diblock copolymer which has the least effect on the fluorescence properties of the FPs. According to the present results, the ideal partners of an FRET-pair in C3Ms would be mTurquoise2 and SYFP2 in combination with P2MVP128-b-PEO477.
4. Materials and Methods
4.1. Materials
Poly(2-vinyl-pyridinium)
n-
b-poly(ethylene-oxide)
m (P2VP
n-
b-PEO
m) with different chain lengths was quaternized: P2VP
41-
b-PEO
205 (Polymer Source
Inc., Dorval, Quebec, Canada,
Mw/
Mn = 1.05,
Mn = 13.3 kg/mol) and P2VP
128-
b-PEO
477 (Polymer Source
Inc., Canada,
Mw/
Mn = 1.10,
Mn = 34.5 kg/mol), following a procedure described elsewhere [
18]. For P2MVP
41-
b-PEO
205 (
Mn = 18.6 kg/mol) and for P2MVP
128-
b-PEO
477 (
Mn = 50.8 kg/mol) a final degree of quaternization of approximately 80% and 87% was obtained, respectively. Stock solutions of P2MVP
41-
b-PEO
205 (51 μM) and P2MVP
128-
b-PEO
477 (50 μM) were prepared by dissolving the polymers in 10 mM borate buffer (pH 9.0) and stored at −20 °C. All solutions were filtered through 0.20 μm polyethersulfone membrane syringe filters (Advanced Microdevices Pvt. Ltd., Ambala Cantt, India). All other chemicals were from commercial sources and of the purest grade available.
4.2. Protein Production
The cDNA’s coding for mTurquoise2, SYFP2-His, and mCherry were cloned into the bacterial expression vector pTYB12 (New England Biolabs, Ipswich, MA, USA) to generate FP fusions with a chitin-binding domain and an intein [
52,
53,
54]. The cDNA’s of SBFP2, mEGFP, mKO2, and TagRFP in pRSETb vectors were kindly provided by Dr. J. Goedhart, University of Amsterdam [
22,
23,
24]. For protein production,
E. coli BL21 cells were used. Details on protein production and purification are described elsewhere [
19]. FPs without the chitin-binding domain were acquired after on-column cleavage of mTurquoise2, SYFP2-His, and mCherry. The other FPs, i.e., SBFP2, mEGFP, mKO2, and TagRFP still contained the His-tag after purification. Purified protein samples were stored in 10 mM borate buffer (pH 9.0) at −20 °C.
Protein concentrations were determined with a Pierce BCA protein assay (Pierce Biotechnology, Rockford, IL, USA), using a bovine serum albumin standard as a reference. The purity of the FPs was checked by SDS-PAGE.
4.3. Modeling
Homology models were built from existing crystal structures using SWISS-MODEL [
55,
56,
57,
58].
Table 3 shows the proteins used in this paper and their corresponding PDBs.
Table 4 shows the proteins used in this paper and their respective templates used for the homology modeling. The chromophores were placed in the model structure at the same position and orientation as the chromophore in the template structure. Pairwise sequence alignments of the FPs are listed in
Figures S7–S13. Because some N- and C-termini were missing in the created homology models (for SBFP2, mEGFP, and SYFP2), these termini were modeled manually using the PDB entry 3ZTF as a template. The A206K mutants were created by mutagenesis of Ala206 into Lys206 in PDB entries 4EUL and 3ZTF to construct mEGFP and mTurquoise2, respectively.
Table 3.
PDB structures used for the proteins studied in this research listed with their corresponding figures, sequence identities (% ID) and references.
Table 3.
PDB structures used for the proteins studied in this research listed with their corresponding figures, sequence identities (% ID) and references.
| Protein | PDB Entry | Figure | % ID | Reference |
|---|
| mTurquoise2 | 3ZTF | S8 | 99.57 | Goedhart, et al. [23] |
| mEGFP | 4EUL | S9 | 99.56 | Arpino, et al. [29] |
| TagRFP | 3M22 | S12 | 100.00 | Subach, et al. [59] |
| mCherry | 2H5Q | S13 | 100.00 | Shu, et al. [27] |
Table 4.
Homology models built from PDB entry templates for the proteins studied in this research listed with their corresponding figures, sequence identities (% ID) and references.
Table 4.
Homology models built from PDB entry templates for the proteins studied in this research listed with their corresponding figures, sequence identities (% ID) and references.
| Protein | PDB Entry | Figure | % ID | Reference |
|---|
| SBFP2 | 1BFP | S7 | 96.44 | Wachter, et al. [60] |
| SYFP2 | 1MYW | S10 | 99.12 | Rekas, et al. [61] |
| mKO2 | 2ZMU | S11 | 95.31 | Kikuchi, et al. [62] |
4.4. C3M Preparation
Encapsulation of FPs with polymers was achieved by first diluting the FP stock solution in 10 mM borate buffer at pH 9.0 for SBFP2, mTurquoise2, mEGFP, SYFP2, mKO2, and mCherry; and at pH 10.0 for TagRFP to the desired concentration, followed by addition of the polymer. After mixing, samples were stored at room temperature for 24 h before measuring. All experiments were performed in 10 mM borate buffer at the encapsulation pH.
4.5. Dynamic Light Scattering (DLS)
DLS measurements were performed on an ALV instrument equipped with a 300 mW Cobolt Samba-300 DPSS laser operating at 660 nm and 100 mW, and static and dynamic enhancer fiber optics for an ALV/High QE APD (high quantum efficiency avalanche photo diode) single photon detector connected to an ALV5000/60X0 External Correlator (ALV-Laser Vertriebsgesellschaft m-b.H., Langen, Germany). The detection angle θ was set at 90° and all measurement were performed at room temperature.
DLS measures fluctuations in scattered light intensities caused by the diffusion of particles. The diffusion time of particles is dependent on their size: proteins diffuse faster than the encapsulated proteins in C3Ms. Furthermore, larger particles scatter more light, because the scattered light intensity is proportional to
R6, where
R is the particle radius. The formation of more C3Ms leads to higher light intensities, which results in a maximum in the scattered light intensity versus composition plot (
I vs.
F+). The composition at the maximum in scattered light intensity is denoted as the preferred micellar composition (PMC). For determination of the PMC, 500 μL solutions with different polymer/protein compositions were prepared. The protein concentration was kept constant at 1 μM for each composition. The amount of P2MVP
41-
b-PEO
205 or P2MVP
128-
b-PEO
477 was varied to obtain the desired values of
F+:
where [
n+] =
c+N+ refers to the total concentration of positively charged groups on the polymer and [
n−] =
c−N− is the total concentration of negatively charged groups on the protein molecules. The number of charged groups on the diblock copolymer (
N+) taking the degree of quaternization into account, is +33.1 for P2MVP
41-
b-PEO
205 and +112.0 for P2MVP
128-
b-PEO
477, which is used to calculate [
n+]. The net charge of the proteins as a function of pH was calculated using the software package PROPKA 3.1 [
32,
33]. The charges of the native proteins at pH 9 or 10 (
N−) are listed in
Table 1, which are used to calculate [
n−].
4.6. DLS Data Analysis
DLS autocorrelation curves were generated from 10 intensity traces and averaged. The CUMULANT method [
63,
64] was used to analyze the mean apparent hydrodynamic radius (
Rh) as:
where
q is the scattering vector,
k is the Boltzmann constant,
T is the absolute temperature,
n is the viscosity of the solvent, and
Γ is the measured average decay rate of the correlation function. The CONTIN method [
65,
66] is used to analyze the distribution of the radii of the C3Ms. The data were analyzed with the AfterALV program (AfterALV 1.0d, Dullware, The Netherlands).
4.7. Fluorescence Correlation Spectroscopy (FCS)
FCS was performed on a Leica TCS SP8 X SMD system equipped with a 63× 1.20 NA (numeric aperture) water immersion objective with coverslip thickness correction collar. Samples with FPs were excited using a diode laser (emits at 440 nm) or a super continuum laser (emits a continuous spectrum from 470 to 670 nm). The lasers were set at a pulsed frequency of 40 MHz. The size-adjustable pinhole was set at 70 μm for all measurements. Fluorescence emission was detected using bandpass-adjustable spectral filters. In
Table 5 the used laser lines and range of the spectral filters are given per fluorescent protein. Fluorescence was recorded via the internal hybrid detector, which was coupled to a PicoHarp 300 TCSPC module (PicoQuant, Berlin, Germany). With this system, it was not possible to measure SBFP2, because its excitation maximum is below 440 nm.
Rhodamine 110 (D = 4.3 × 10−10 m2 s−1) was used to calibrate the confocal volume of the setup. A diffusion time of 18 μs and a structural parameter (a, expressed as (ωxy/ωz)) between 5 and 10 were obtained, resulting in a confocal volume of approximately 0.2 fL. Measurements were performed in a μ-Slide 8-wells chambered coverslip (Ibidi®).
Samples with concentrations of 1 μM FP were measured free in buffered solution as well as encapsulated with P2MVP41-b-PEO205 or P2MVP128-b-PEO477 at their respective PMCs. For each sample, 5 fluorescence intensity fluctuation traces of 30 s each were collected. All measurements were performed at room temperature.
4.8. FCS Data Analysis
For the FCS data analysis, the FFS-data processor version 2.3 (Scientific Software Technologies Software Centre, Minsk, Belarus) was used [
67]. The equation used to fit translational data, which includes triplet state, is as follows [
68]:
where
represents the average number of fluorescent particles in the confocal volume. The exponential term describes the triplet state behavior of the molecule, in which
Ftrip is the fraction of molecules in the triplet state and
Ttrip is the average time a molecule resides in the triplet state. The last part of the equation describes the diffusion behavior of the molecules, where
Fi is the fraction of species
i, τ
diff,i is the diffusion time of species
i, ω
xy and ω
z are the equatorial and axial radii of the detection volume, respectively. Equation (3) was used to obtain
for the different samples.
4.9. Steady-State Fluorescence Spectroscopy
Fluorescence excitation and emission spectra were measured using a Cary Eclipse spectrofluorimeter (Varian). Excitation and emission slits were set to yield bandwidths of 5 nm. All measurements were performed at 20 °C. Samples with concentrations of 1 μM FP were measured free in buffered solution as well as encapsulated with P2MVP41-b-PEO205 or P2MVP128-b-PEO477 at their respective PMCs.
The relative quantum yields are calculated using the following equation [
69]:
where QY represents the quantum yield, FA the integrated area under the corrected emission spectrum, and A the absorbance at the excitation wavelength. The subscripts
C3M and
P refer to the proteins in the C3Ms and the proteins free in solution, respectively.
4.10. Circular Dichroism (CD)
CD experiments were performed on a JASCO J-715 spectropolarimeter with a Jasco PTC 348 WI temperature controller set at 20 °C. The far-UV CD spectra (195–260 nm) were obtained from samples in a 0.3 mL quartz cuvette with an optical path length of 1 mm. Thirty spectra, each recorded with a resolution of 1 nm, a scan speed of 50 nm/min and a response time of 1 s, were accumulated and averaged. Samples with concentrations of 2.5 μM FP were measured free in buffered solution as well as encapsulated with P2MVP41-b-PEO205 or P2MVP128-b-PEO477 at their respective PMC. The polymers did not show any CD signal over the measured range, therefore, buffer blank spectra, obtained at identical conditions, were subtracted.
5. Conclusions
We have studied the encapsulation efficiency of SBFP2, mTurquoise2, mEGFP, SYFP2, mKO2, TagRFP, and mCherry and determined their spectral and structural properties as protein free in solution and upon encapsulation with P2MVP41-b-PEO205 or P2MVP128-b-PEO477. This revealed that avFPs are almost 100% encapsulated, while anFPs show encapsulation efficiencies ranging between 50% and 75%. Upon encapsulation, all FPs show differences in spectral properties compared to their respective protein free in solution: the chromophores of SBFP2, mKO2, and mCherry are affected in their molar extinction coefficient and the chromophores of mTurquoise2, mEGFP, SYFP2, TagRFP, and mKO2 are affected in their fluorescence quantum yield. Only for mCherry, the changes in spectral properties upon encapsulation are similar to changes observed as a result of pH variation and are, therefore, related to a shift in the pKa. Even though all FPs have an 11-stranded β-barrel fold, the CD spectra differ between avFPs and anFPs. This is most likely due to a different shape of the cylinders between the two groups of FPs, where the β-barrel structures of avFPs are almost round cylinders and that of anFPs elliptic ones. This variation in structure, together with the difference in charge distribution on FP surfaces, potentially causes the differences in encapsulation efficiency.

 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}







