A Strategy to Replace the Mouse Bioassay for Detecting and Identifying Lipophilic Marine Biotoxins by Combining the Neuro-2a Bioassay and LC-MS/MS Analysis

 ,
,
Abstract
:1. Introduction
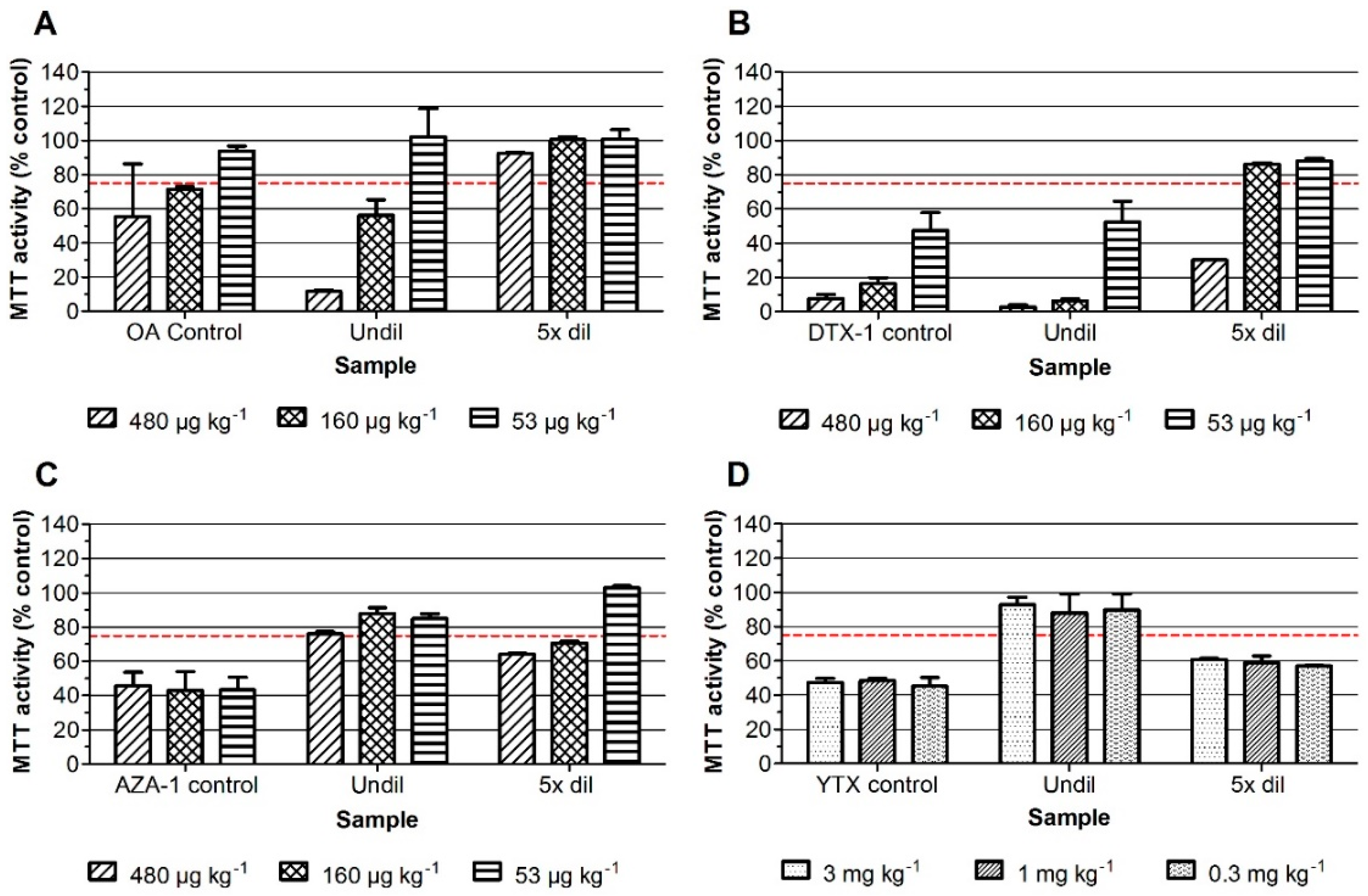
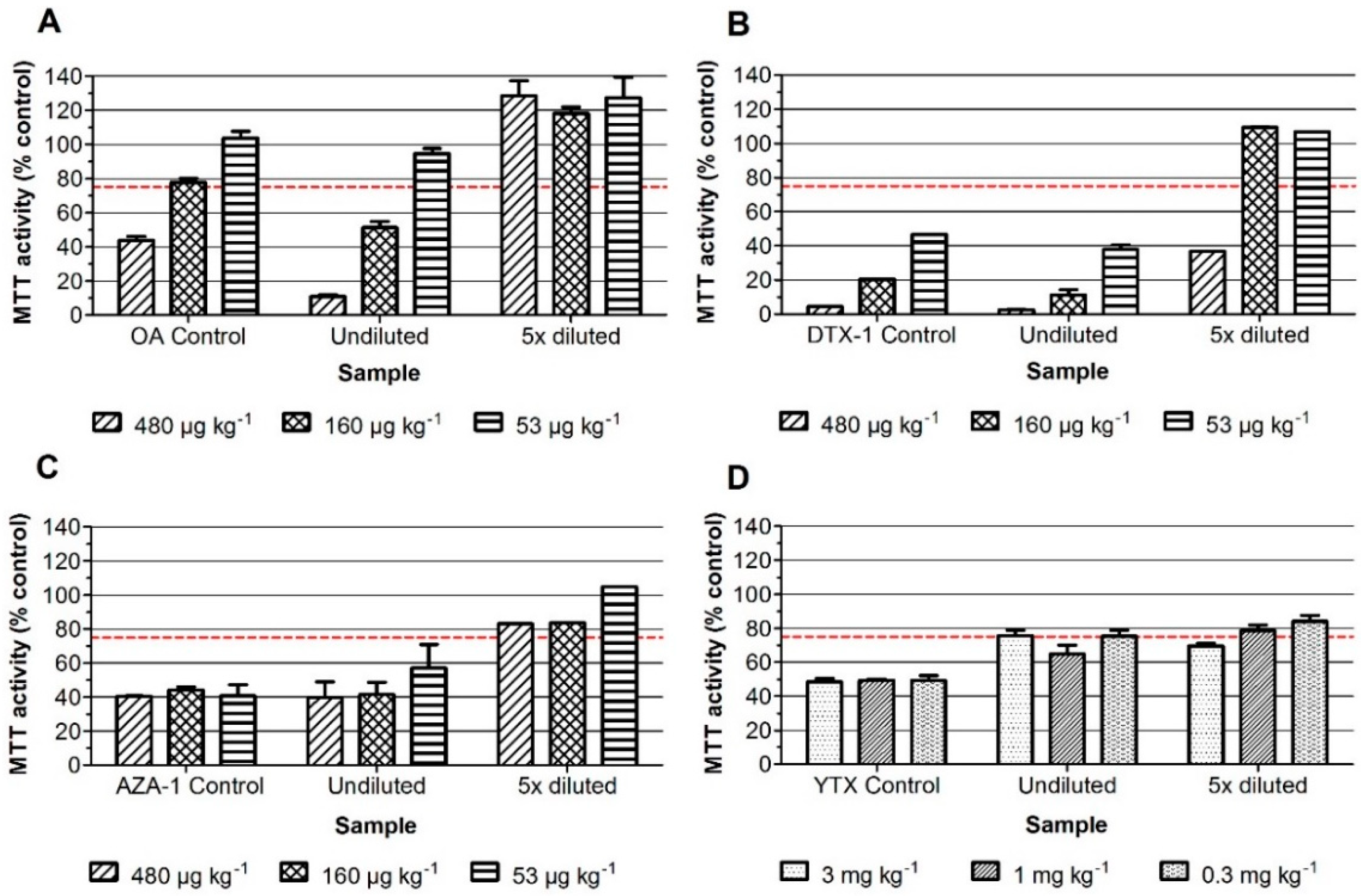
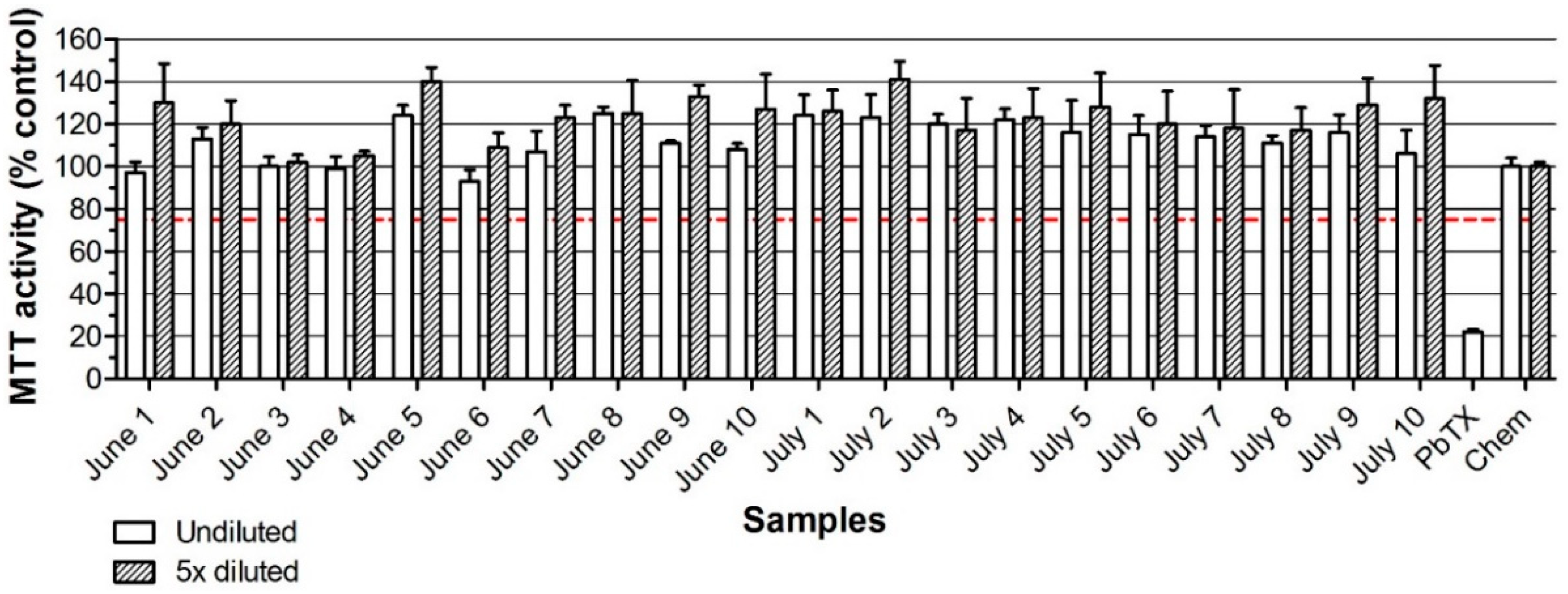
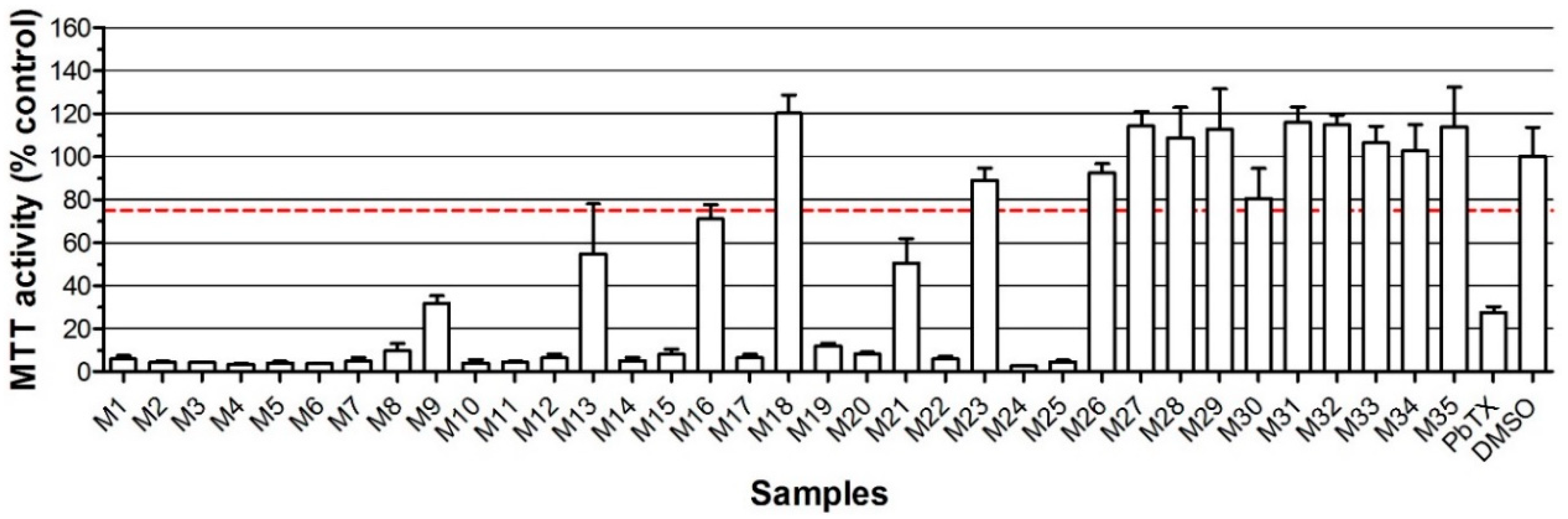
2. Results
2.1. Neuro-2a Bioassay with Mussel Samples
2.2. Neuro-2a Bioassay with Naturally Contaminated Samples Tested Previously in the MBA
3. Discussion
4. Materials and Methods
4.1. Reagents and Standards
4.2. Samples
4.3. Sample Extraction
4.4. Further Sample Clean-Up by Washing with n-Hexane Followed by SPE
4.5. Fortification of Samples
4.6. Neuro-2a Bioassay
4.7. LC-MS/MS Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Zamorano, R.; Marín, M.; Cabrera, F.; Figueroa, D.; Contreras, C.; Barriga, A.; Lagos, N.; García, C. Determination of the variability of both hydrophilic and lipophilic toxins in endemic wild bivalves and carnivorous gastropods from the Southern part of Chile. Food Addit. Contam. Part A-Chem. 2013, 30, 1660–1677. [Google Scholar] [CrossRef] [PubMed]
- Stewart, I.; McLeod, C. The laboratory mouse in routine food safety testing for marine algal biotoxins and harmful algal bloom toxin research: Past, present and future. J. AOAC Int. 2014, 97, 356–372. [Google Scholar] [CrossRef] [PubMed]
- Visciano, P.; Schirone, M.; Berti, M.; Milandri, A.; Tofalo, R.; Suzzi, G. Marine Biotoxins: Occurrence, Toxicity, Regulatory Limits and Reference Methods. Frontiers Microb. 2016, 7, 1051. [Google Scholar] [CrossRef] [PubMed]
- Rossini, G.P. Functional assays in marine biotoxin detection. Toxicology 2005, 207, 451–462. [Google Scholar] [CrossRef] [PubMed]
- Bodero, M.; Bovee, T.F.H.; Wang, S.; Hoogenboom, R.; Klijnstra, M.D.; Portier, L.; Hendriksen, P.J.M.; Gerssen, A. Screening for the presence of lipophilic marine biotoxins in shellfish samples using the neuro–2a bioassay. Food Addit. Contam. Part A-Chem. 2018, 35, 351–365. [Google Scholar] [CrossRef] [PubMed]
- Cañete, E.; Diogène, J. Improvements in the use of neuroblastomaxglioma hybrid cells (NG108–15) for the toxic effect quantification of marine toxins. Toxicon 2010, 55, 381–389. [Google Scholar] [CrossRef] [PubMed]
- Nicolas, J.; Hendriksen, P.J.; Gerssen, A.; Bovee, T.F.; Rietjens, I.M. Marine neurotoxins: State of the art, bottlenecks, and perspectives for mode of action based methods of detection in seafood. Mol. Nutr. Food Res. 2014, 58, 87–100. [Google Scholar] [CrossRef] [PubMed]
- Reverté, L.; Solino, L.; Carnicer, O.; Diogène, J.; Campàs, M. Alternative methods for the detection of emerging marine toxins: Biosensors, biochemical assays and cell-based assays. Mar. Drugs 2014, 12, 5719–5763. [Google Scholar] [CrossRef] [PubMed]
- European Commission. Commission Regulation (EU) No 15/2011 of 10 January 2011 amending Regulation (EC) No 2074/2005 as regards recognised testing methods for detecting marine biotoxins in live bivalve molluscs. Off. J. Eur. Commun. 2011, 50, 3–4. [Google Scholar]
- Toyofuku, H. Joint FAO/WHO/IOC activities to provide scientific advice on marine biotoxins (research report). Mar. Pollut. Bull. 2006, 52, 1735–1745. [Google Scholar] [CrossRef] [PubMed]
- Paredes, I.; Rietjens, I.M.; Vieites, J.M.; Cabado, A.G. Update of risk assessments of main marine biotoxins in the European Union. Toxicon 2011, 58, 336–354. [Google Scholar] [CrossRef] [PubMed]
- Lombet, A.; Bidard, J.N.; Lazdunski, M. Ciguatoxin and brevetoxins share a common receptor site on the neuronal voltage-dependent Na+ channel. FEBS Lett. 1987, 219, 355–359. [Google Scholar] [CrossRef]
- Todd, K. A Review of NSP Monitoring in New Zealand in Support of a New Programme; Cawthron Institute: Nelson, New Zealand, 2002. [Google Scholar]
- Terzagian, R. Five Cluster of Neurotoxic Shellfish Poisoning (NSP) in Lee Country; Florida Department of Health: Florida, FL, USA, July 2006. [Google Scholar]
- Dechraoui, M.Y.; Tiedeken, J.A.; Persad, R.; Wang, Z.; Granade, H.R.; Dickey, R.W.; Ramsdell, J.S. Use of two detection methods to discriminate ciguatoxins from brevetoxins: Application to great barracuda from Florida Keys. Toxicon 2005, 46, 261–270. [Google Scholar] [CrossRef] [PubMed]
- Denisson, N.; Anderson, D.B. The 3 “R” S approach to marine biotoxin testing in the UK. In Proceedings of the 6th World Congress on Alternatives and Animal Use in the Life Sciences, Tokyo, Japan, 21–25 August 2007; pp. 757–761. [Google Scholar]
- Hess, P.; Grune, B.; Anderson, D.B.; Aune, T.; Botana, L.M.; Caricato, P.; van Egmond, H.P.; Halder, M.; Hall, S.; Lawrence, J.F.; et al. Three Rs Approaches in Marine Biotoxin Testing. The Report and Recommendations of a joint ECVAM/DG SANCO Workshop (ECVAM Workshop 54). Alter. Lab. Animals ATLA 2006, 34, 193–224. [Google Scholar]
- Campbell, K.; Vilariño, N.; Botana, L.M.; Elliott, C.T. A European perspective on progress in moving away from the mouse bioassay for marine-toxin analysis. Trends Analy. Chem. 2011, 30, 239–253. [Google Scholar] [CrossRef]
- Botana, L.M.; Vilariño, N.; Alfonso, A.; Vale, C.; Louzao, C.; Elliott, C.T.; Campbell, K.; Botana, A.M. The problem of toxicity equivalent factors in developing alternative methods to animal bioassays for marine-toxin detection. Trends Analy. Chem. 2010, 29, 1316–1325. [Google Scholar] [CrossRef]
- EFSA. Scientific Opinion of the Panel on Contaminants in the Food Chain on a request from the European Commission on Marine Biotoxins in Shellfish – Summary on regulated marine biotoxins. EFSA J. 2009, 1306, 1–23. [Google Scholar]
- Buckland, G. Implementing scientifically-robust and humane shellfish toxicity testing: We’re still waiting. Alter. Lab. Animals ATLA 2010, 38, 419–426. [Google Scholar]
- Botana, L.M. A perspective on the toxicology of marine toxins. Chem. Res. Toxicol. 2012, 25, 1800–1804. [Google Scholar] [CrossRef]
- Cañete, E.; Diogène, J. Comparative study of the use of neuroblastoma cells (Neuro–2a) and neuroblastoma × glioma hybrid cells (NG108–15) for the toxic effect quantification of marine toxins. Toxicon 2008, 52, 541–550. [Google Scholar] [CrossRef]
- Manger, R.L.; Leja, L.S.; Lee, S.Y.; Hungerford, J.M.; Hokama, Y.; Dickey, R.W.; Granade, H.R.; Lewis, R.; Yasumoto, T.; Wekell, M.M. Detection of sodium channel toxins: Directed cytotoxicity assays of purified ciguatoxins, brevetoxins, saxitoxins, and seafood extracts. J. AOAC Int. 1995, 78, 521–527. [Google Scholar] [PubMed]
- European Commission. Commission Decision (EC) No 2002/657, Commission Decision of 12 August 2002 implementing Council Directive 96/23/EC concerning the performance of analytical methods and the interpretation of results (Text with EEA relevance) (notified under document number C(2002) 3044). Off. J. Eur. Commun. 2002, 8–36. [Google Scholar]
- Bodero, M.; Bovee, T.F.H.; Portier, L.; Hendriksen, P. Detection and profiling of diarroeic marine biotoxins in shellfish by mRNA analysis of exposed Caco-2 cells using qRT-PCR and multiplex magnetic bead-based assay. ALTEX 2018, in press. [Google Scholar]
- Yasumoto, T.; Murata, M.; Oshima, Y.; Matsumoto, G.K.; Clardy, J.O.N. Diarrhetic Shellfish Poisoning. Seafood Toxins 1984, 262, 207–214. [Google Scholar]
- Gerssen, A.; van Olst, E.H.; Mulder, P.P.; de Boer, J. In-house validation of a liquid chromatography tandem mass spectrometry method for the analysis of lipophilic marine toxins in shellfish using matrix–matched calibration. Anal. Bioanal. Chem. 2010, 397, 3079–3088. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Species | MBA | Neuro 2a with o/v | Neuro2a without o/v | LC-MS/MS (µg kg−1) | ||
|---|---|---|---|---|---|---|---|
| OA eq | AZA-1 eq | YTX eq | |||||
| M1 | Mussel | positive | + | + | 898 | ||
| M2 | Mussel | positive | + | + | 869 | ||
| M3 | Mussel | ND | + | + | 704 | ||
| M4 | Mussel | positive | + | + | 1060 | ||
| M5 | Mussel | positive | + | + | 569 | ||
| M6 | Clam | positive | + | + | |||
| M7 | Clam | ND | + | + | |||
| M8 | Clam | positive | + | + | |||
| M9 | Mussel | positive | + | + | 1459 | ||
| M10 | Mussel | positive | + | + | 2531 | ||
| M11 | Clam | ND | + | + | |||
| M12 | Mussel | positive | + | + | 1648 | ||
| M13 | Mussel | positive | + | + | 1271 | ||
| M14 | Mussel | positive | + | + | 22 | 1546 | |
| M15 | Mussel | positive | + | + | 1254 | ||
| M16 | Clam | ND | + | - | |||
| M17 | Clam | positive | + | + | |||
| M18 | Clam | ND | - | - | |||
| M19 | Mussel | positive | + | + | 33 | 1953 | |
| M20 | Mussel | positive | + | + | 21 | 1432 | |
| M21 | Mussel | positive | + | + | 20 | 1518 | |
| M22 | Mussel | positive | + | + | 1141 | ||
| M23 | Clam | ND | - | + | |||
| M24 | Mussel | positive | + | + | 1067 | ||
| M25 | Mussel | positive | + | + | 1648 | ||
| M26 | Mussel | ND | - | + | 272 | ||
| M27 | Mussel | ND | - | - | |||
| M28 | Clam | ND | - | - | |||
| M29 | Mussel | ND | - | - | |||
| M30 | Clam | ND | - | - | 15 | ||
| M31 | Mussel | ND | - | - | |||
| M32 | Clam | ND | - | - | 9 | ||
| M33 | Mussel | ND | - | - | |||
| M34 | Mussel | ND | - | - | |||
| M35 | Mussel | ND | - | - | |||
| Compound Name | Precursor Ion (m/z) | Product Ion (m/z) | Ionization Polarity | DP 1 (V) | CE 2 (V) | CXP 3 (V) | Standard Available |
|---|---|---|---|---|---|---|---|
| Yessotoxin | 570.4 | 396.4 | Negative | −45 | −42 | −17 | Yes |
| 570.4 | 467.4 | Negative | −45 | −42 | −13 | ||
| Homo yessotoxin | 577.4 | 403.4 | Negative | −45 | −42 | −17 | Yes |
| 577.4 | 474.4 | Negative | −45 | −42 | −13 | ||
| 45OH yessotoxin | 578.4 | 396.4 | Negative | −45 | −42 | −17 | No |
| 578.4 | 467.4 | Negative | −45 | −42 | −13 | ||
| 45OH Homo yessotoxin | 585.4 | 403.4 | Negative | −45 | −42 | −17 | No |
| 585.4 | 474.4 | Negative | −45 | −42 | −13 | ||
| Okadaic acid & dinophysistoxin-2 | 803.5 | 563.1 | Negative | −15 | −62 | −19 | Yes |
| 803.5 | 255.2 | Negative | −15 | −60 | −55 | ||
| Dinophysistoxin-1 | 817.5 | 563.2 | Negative | −15 | −62 | −19 | Yes |
| 817.5 | 255.2 | Negative | −15 | −60 | −55 | ||
| Gymnodimine | 508.2 | 120.0 | Positive | 66 | 107 | 18 | Yes |
| 508.2 | 490.2 | Positive | 66 | 35 | 36 | ||
| 13 desmethyl spirolide C | 692.5 | 164.3 | Positive | 40 | 40 | 12 | Yes |
| 692.5 | 444.2 | Positive | 40 | 55 | 32 | ||
| Pinnatoxin-G | 694.5 | 164.3 | Positive | 40 | 63 | 10 | Yes |
| 694.5 | 676.5 | Positive | 40 | 51 | 18 | ||
| 20 methyl spirolide G | 706.5 | 164.3 | Positive | 40 | 40 | 12 | Yes |
| 706.5 | 346.2 | Positive | 40 | 55 | 32 | ||
| Azaspiracid-3 | 828.5 | 654.3 | Positive | 41 | 67 | 18 | Yes |
| 828.5 | 810.5 | Positive | 41 | 43 | 22 | ||
| Azaspiracid-1 | 842.5 | 672.4 | Positive | 41 | 67 | 18 | Yes |
| 842.5 | 824.5 | Positive | 41 | 43 | 22 | ||
| Azaspiracid-2 | 856.5 | 672.4 | Positive | 41 | 67 | 18 | Yes |
| 856.5 | 838.5 | Positive | 41 | 43 | 22 |
| Compound Name | Precursor Ion (m/z) | Product Ion (m/z) | Ionization Polarity | Cone (V) | Collision (eV) | Standard Available |
|---|---|---|---|---|---|---|
| Brevetoxin 1 | 867.5 | 221.0 | Positive | 40 | 20 | Yes |
| 889.5 | 845.5 | Positive | 40 | 40 | ||
| Brevetoxin 2 | 895.5 | 319.2 | Positive | 40 | 23 | Yes |
| 895.5 | 877.4 | Positive | 40 | 20 | ||
| Brevetoxin 3 | 897.5 | 725.4 | Positive | 40 | 20 | Yes |
| 927.5 | 919.5 | Positive | 40 | 40 | ||
| Brevetoxin 9 | 899.5 | 157.2 | Positive | 40 | 20 | Yes |
| 899.5 | 863.4 | Positive | 40 | 20 | ||
| Brevetoxin B5 | 911.5 | 875.5 | Positive | 40 | 21 | No |
| S-deoxy-brevetoxin-B2 | 1018.6 | 80.96 | Positive | 40 | 80 | No |
| 1018.6 | 204.2 | Positive | 40 | 45 | ||
| Brevetoxin-B2 | 1034.6 | 929.6 | Positive | 40 | 35 | No |
| 1034.6 | 947.5 | Positive | 40 | 35 | ||
| Cys brevetoxin-A | 990.5 | 869.5 | Positive | 40 | 35 | No |
| 990.5 | 901.5 | Positive | 40 | 35 | ||
| Cys brevetoxin-A S-oxide | 1006.5 | 869.5 | Positive | 40 | 35 | No |
| 1006.5 | 919.5 | Positive | 40 | 35 | ||
| Cys brevetoxin-A glycine | 1047.5 | 869.5 | Positive | 40 | 35 | No |
| 1047.5 | 901.5 | Positive | 40 | 35 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bodero, M.; Gerssen, A.; Portier, L.; Klijnstra, M.D.; Hoogenboom, R.L.A.P.; Guzmán, L.; Hendriksen, P.J.M.; Bovee, T.F.H. A Strategy to Replace the Mouse Bioassay for Detecting and Identifying Lipophilic Marine Biotoxins by Combining the Neuro-2a Bioassay and LC-MS/MS Analysis. Mar. Drugs 2018, 16, 501. https://doi.org/10.3390/md16120501
Bodero M, Gerssen A, Portier L, Klijnstra MD, Hoogenboom RLAP, Guzmán L, Hendriksen PJM, Bovee TFH. A Strategy to Replace the Mouse Bioassay for Detecting and Identifying Lipophilic Marine Biotoxins by Combining the Neuro-2a Bioassay and LC-MS/MS Analysis. Marine Drugs. 2018; 16(12):501. https://doi.org/10.3390/md16120501
Chicago/Turabian StyleBodero, Marcia, Arjen Gerssen, Liza Portier, Mirjam D. Klijnstra, Ron L. A. P. Hoogenboom, Leonardo Guzmán, Peter J. M. Hendriksen, and Toine F. H. Bovee. 2018. "A Strategy to Replace the Mouse Bioassay for Detecting and Identifying Lipophilic Marine Biotoxins by Combining the Neuro-2a Bioassay and LC-MS/MS Analysis" Marine Drugs 16, no. 12: 501. https://doi.org/10.3390/md16120501