Offshore Wind Farms as Potential Locations for Flat Oyster (Ostrea edulis) Restoration in the Dutch North Sea

Abstract
:1. Introduction
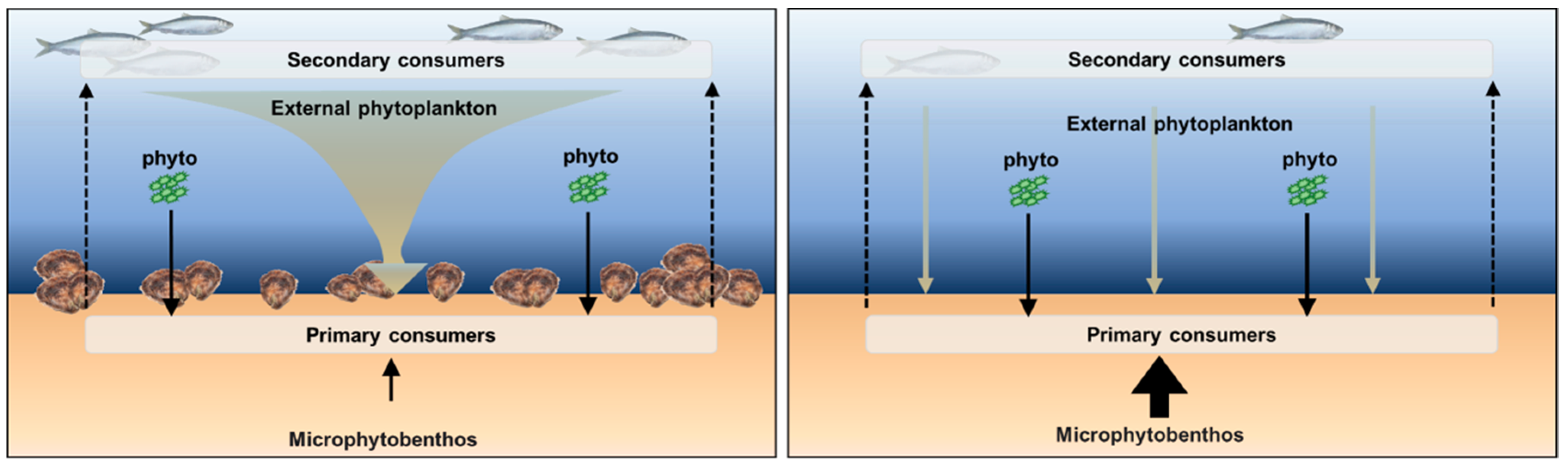
1.1. Flat Oyster Bed Ecosystem Services in the North Sea
1.2. Policy
1.3. Food and Nutrition Security
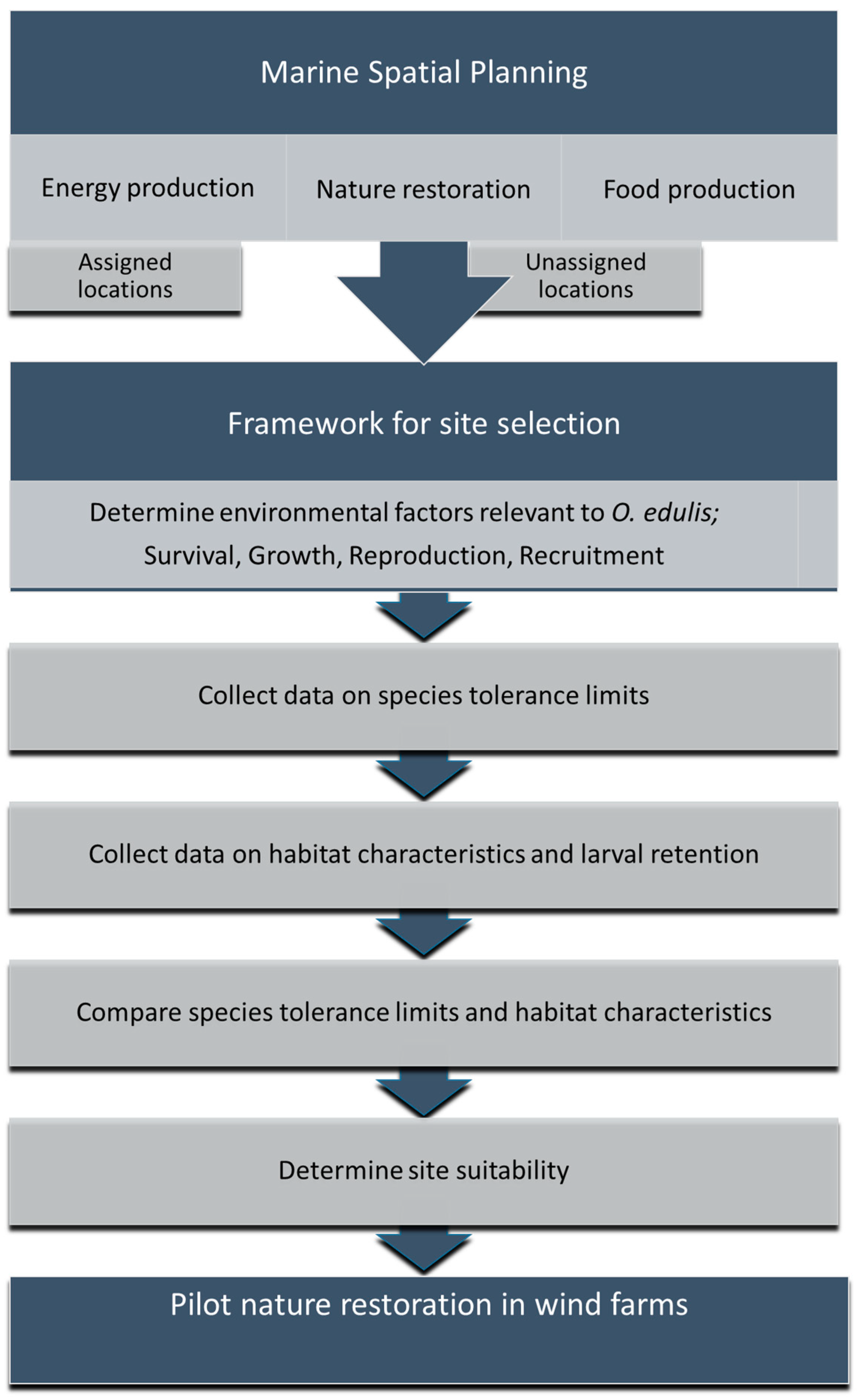
1.4. Wind Farm Suitability
1.5. Life History
2. Materials and Methods
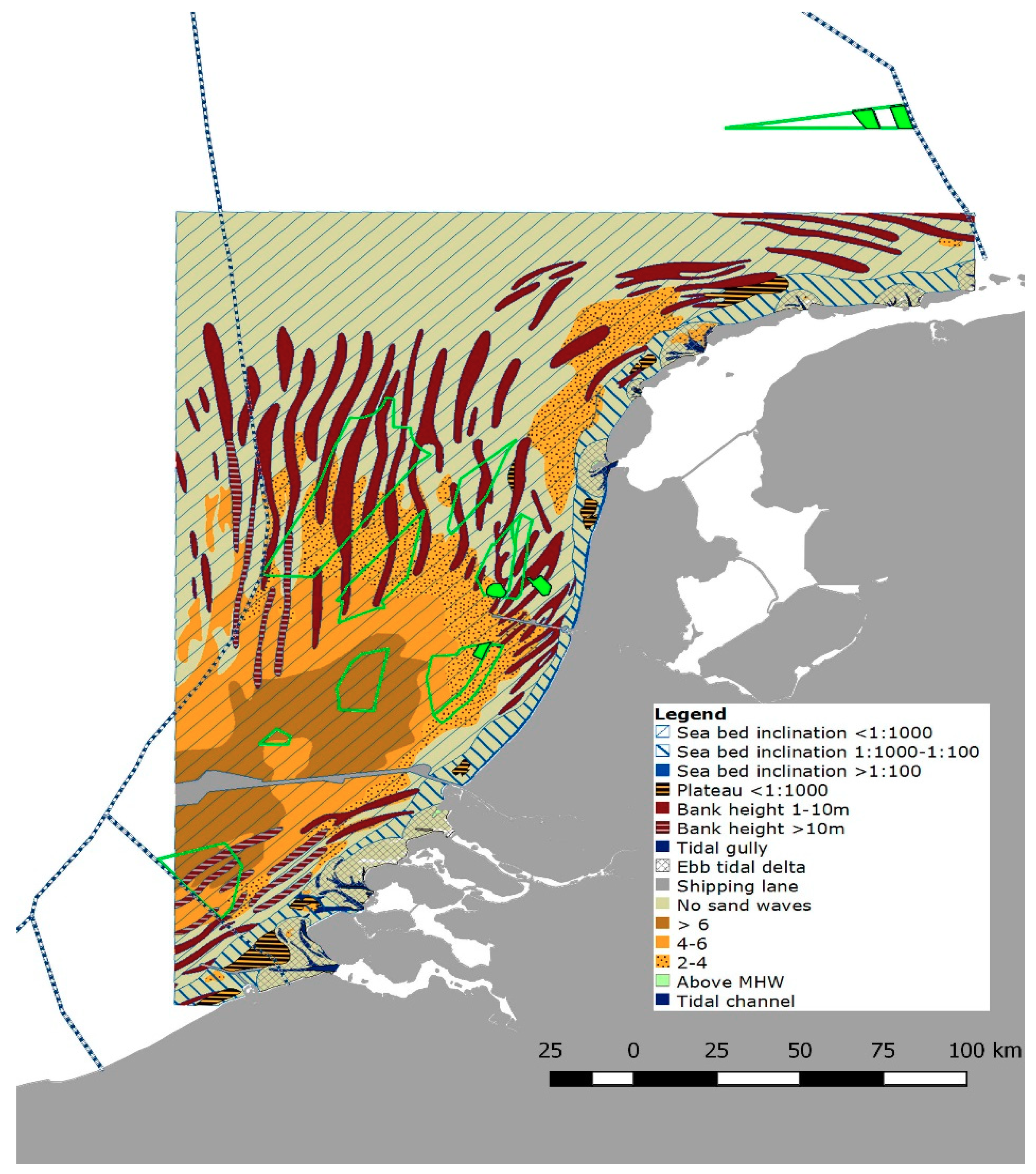
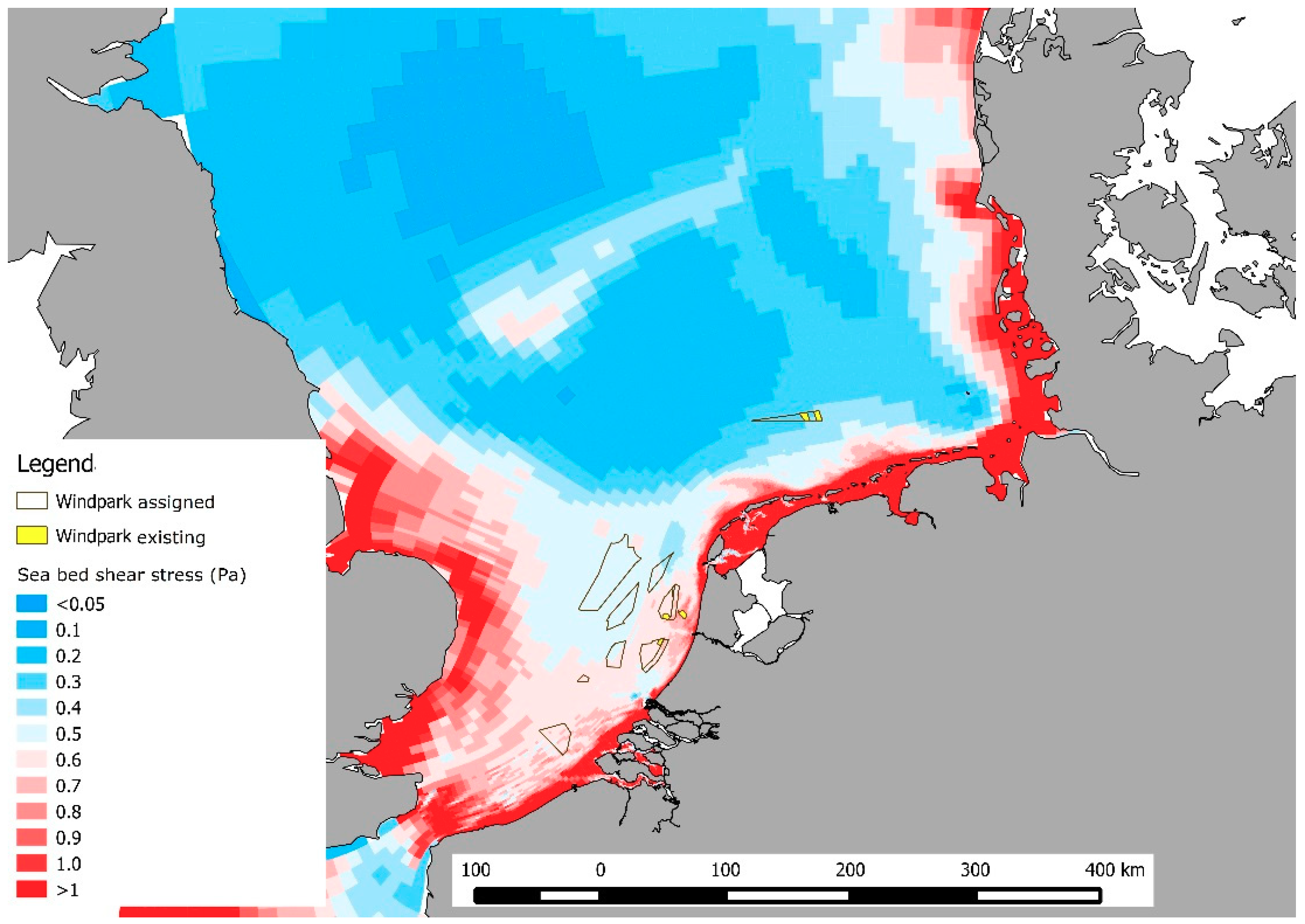
2.1. Sea Bed Motion and Bed Shear Stress
2.2. Concentration of Suspended Particles and Sediment Composition
2.3. Food Availability
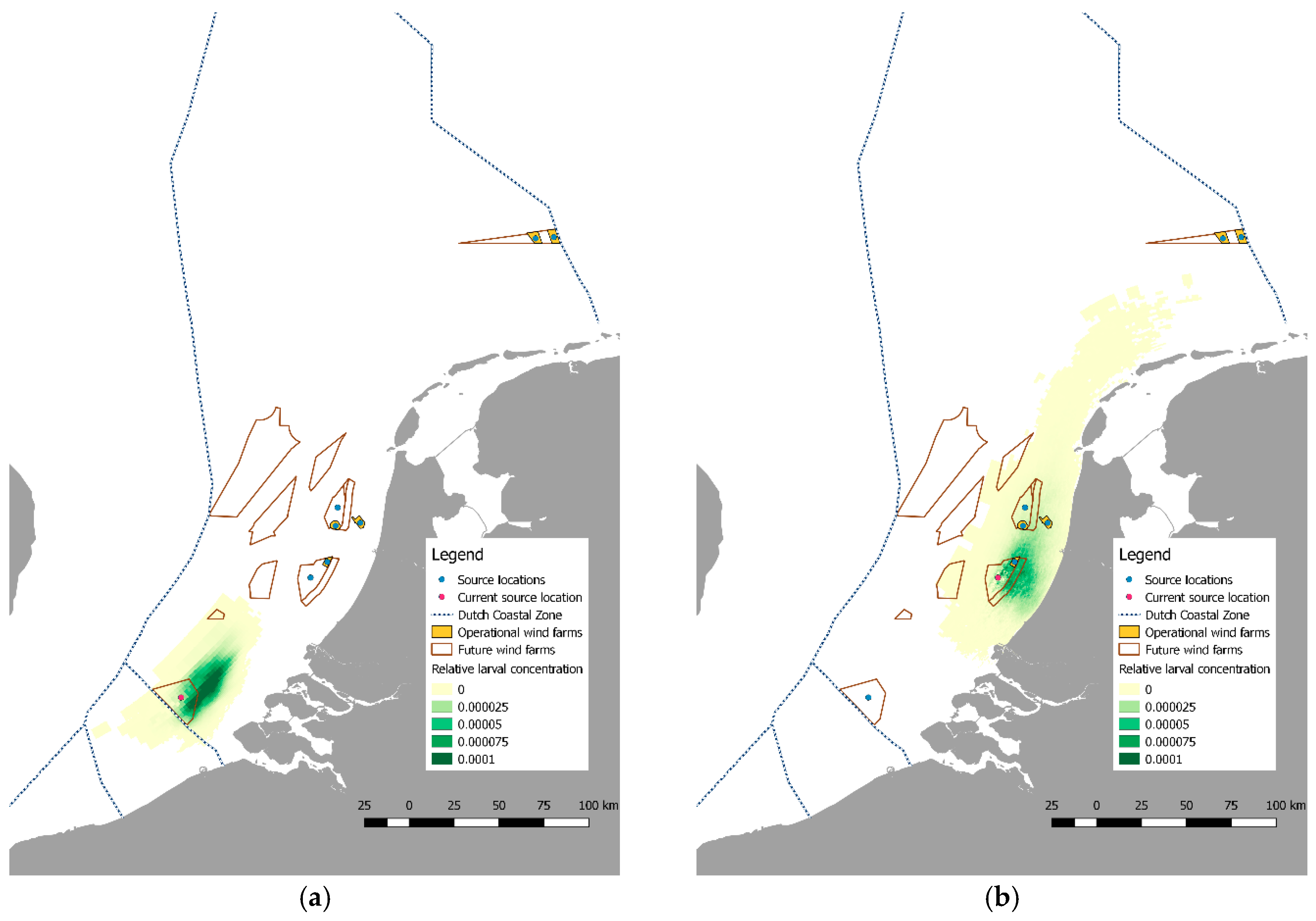
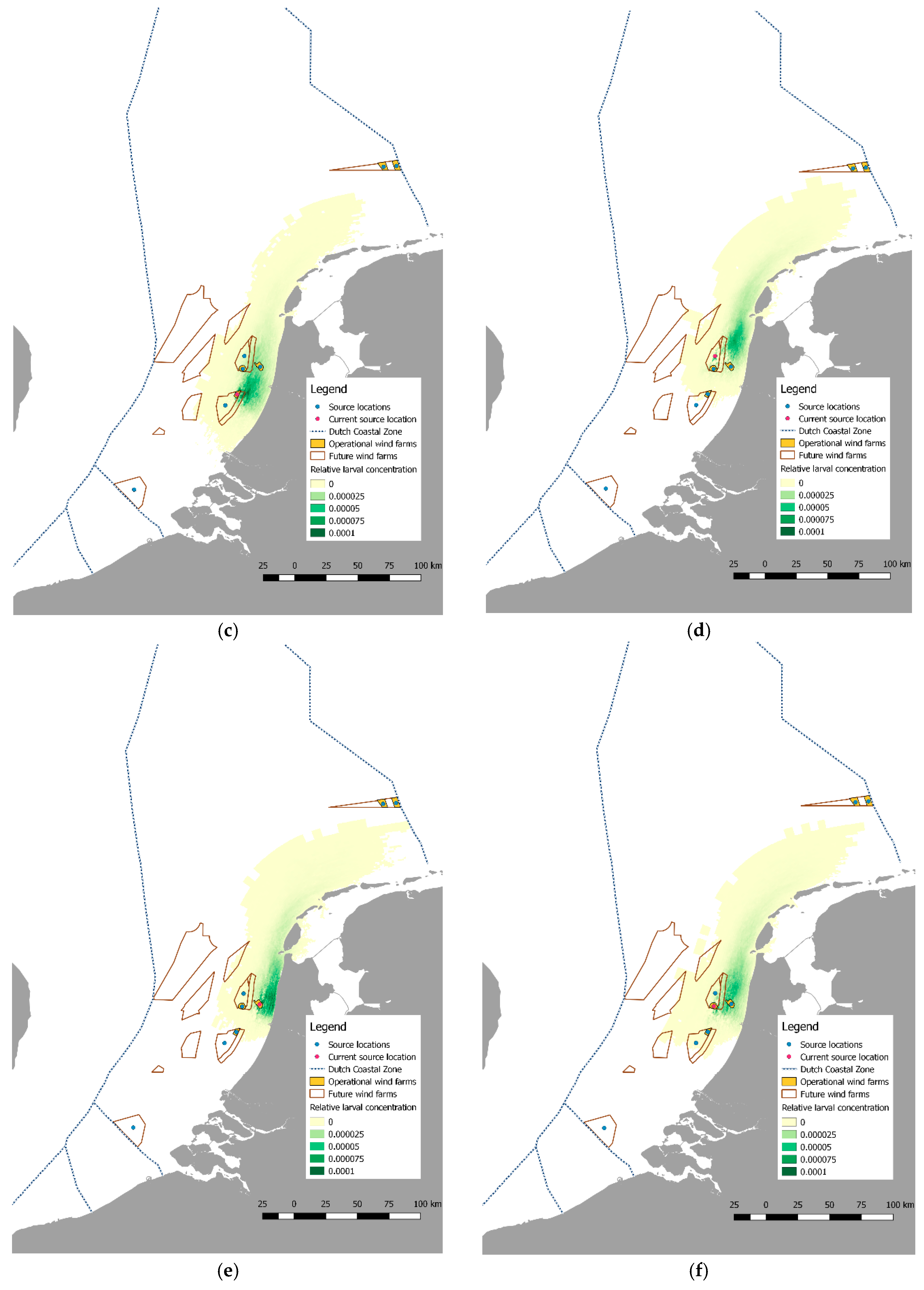
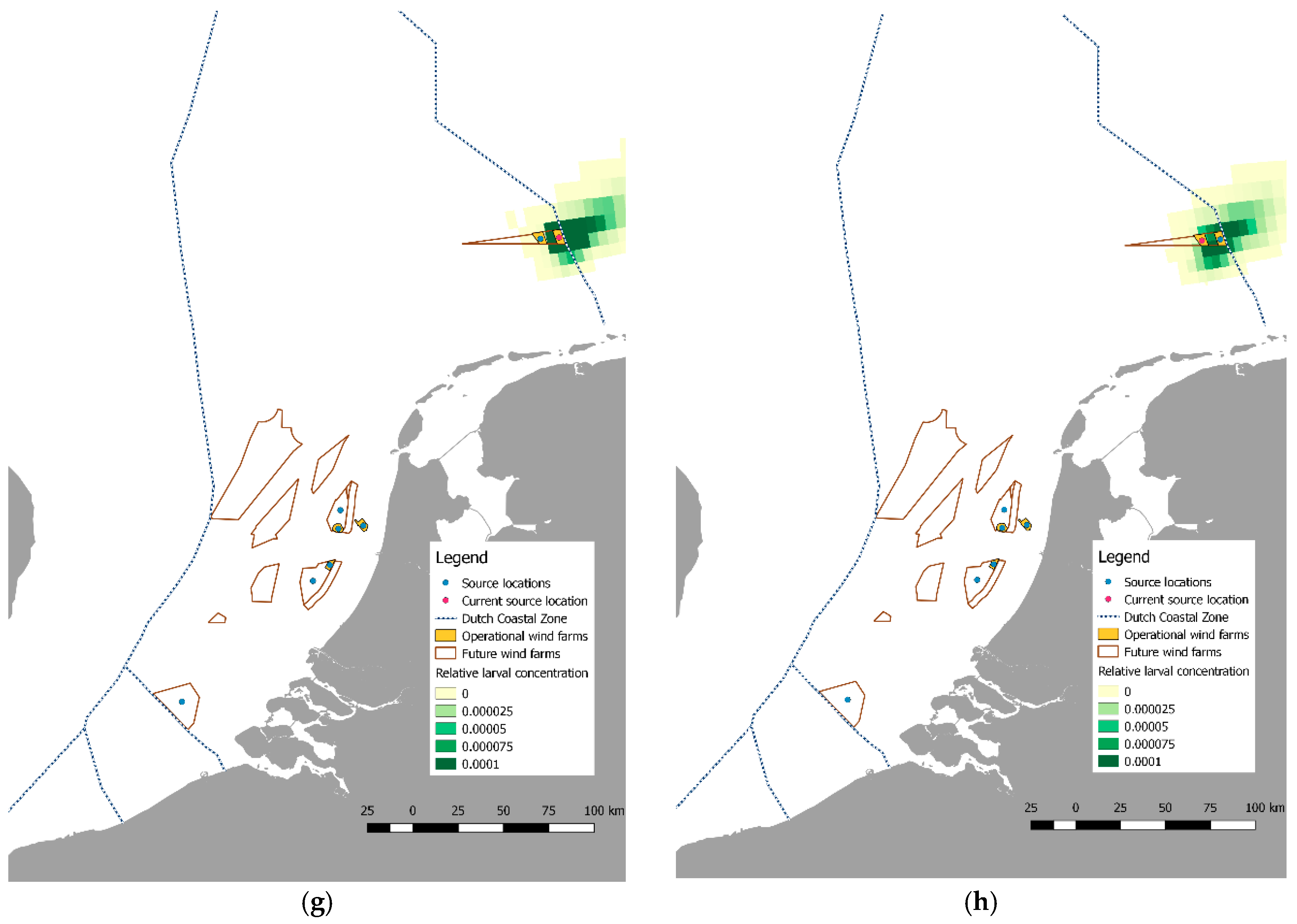
2.4. Larval Dispersal and Retention
2.5. Size of Parent Stock
3. Results and Discussion
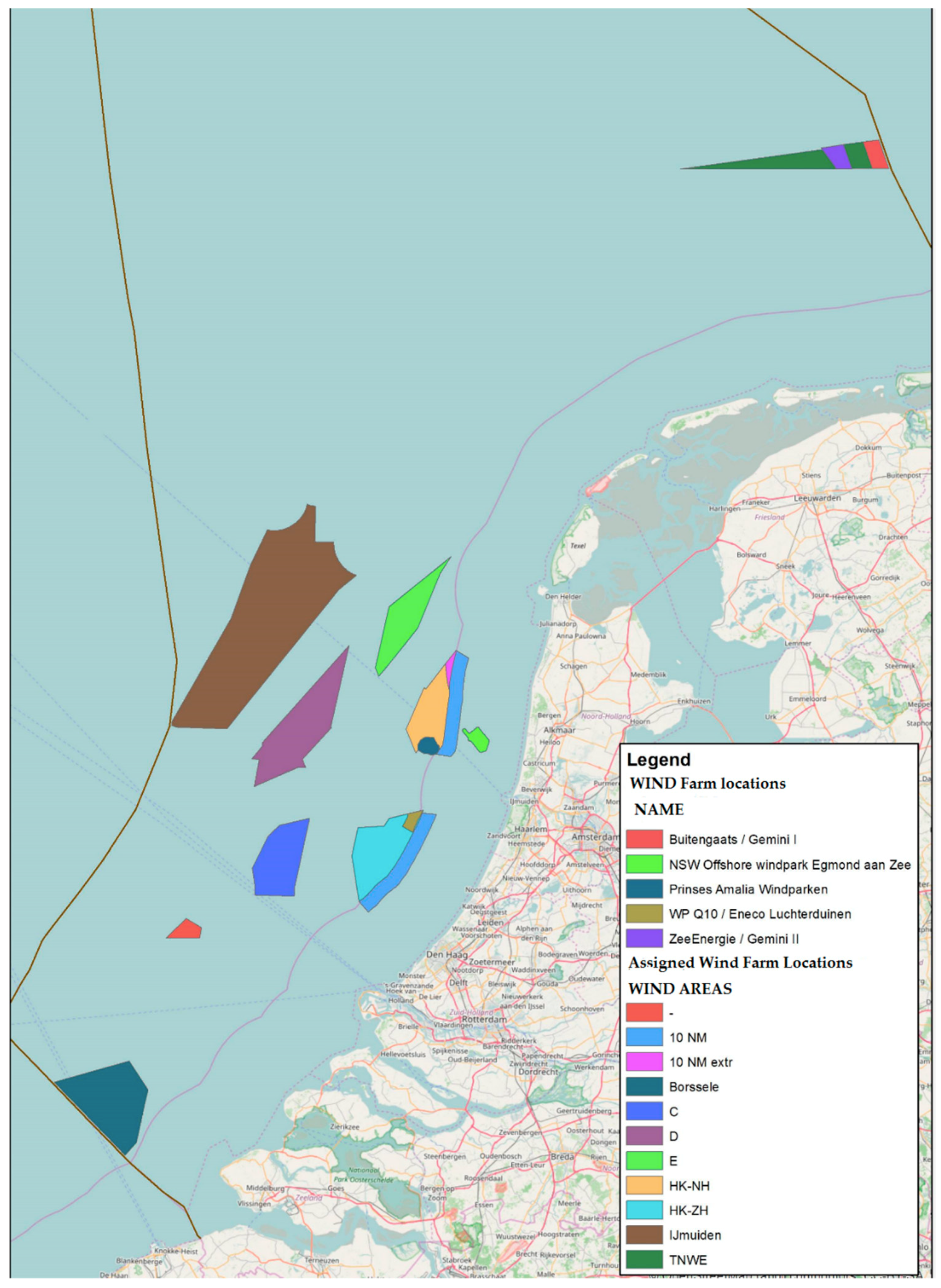
3.1. Wind Farm Selection
3.2. Pilot Studies
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Béné, C.; Barange, M.; Subasinghe, R.; Pinstrup-Andersen, P.; Merino, G.; Hemre, G.; Williams, M. Feeding 9 billion by 2050 Putting fish back on the menu. Food Secur. 2015, 7, 261–274. [Google Scholar] [CrossRef]
- Lovatelli, A.; Aguilar-Manjarrez, J.; Soto, D. (Eds.) Expanding Mariculture Farther Offshore—Technical, Environmental, Spatial and Governance Challenges; FAO Technical Workshop, FAO Fisheries and Aquaculture Proceedings No. 24; Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 2013; p. 73. [Google Scholar]
- Rubino, M.C. Offshore Aquaculture in the United States: Economic Considerations, Implications and Opportunities; Technical Memorandum NMFS F/SPO-103; National Oceanic and Atmospheric Administration: Silver Spring, MD, USA, 2008.
- Godfray, H.C.J.; Beddington, J.R.; Crute, I.R.; Haddad, L.; Lawrence, D.; Muir, J.F.; Pretty, J.; Robinson, S.; Thomas, S.M.; Toulmin, C. Food Security: The Challenge of Feeding 9 Billion People. Science 2010, 327, 812–818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costello, C.; Ovando, D.; Clavelle, T.; Strauss, C.K.; Hilborn, R.; Melnychuk, M.C.; Branch, T.A.; Gaines, S.D.; Szuwalski, C.S.; Cabral, R.B.; et al. Global fishery prospects under contrasting management regimes. PNAS 2016, 113, 5125–5129. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, R.; Naylor, R. Future seascapes, fishing and fish farming. Front. Ecol. Environ. 2005, 3, 21–28. [Google Scholar] [CrossRef]
- Borja, Á.; Rodríguez, J.G.; Black, K.; Bodoy, A.; Emblow, C.; Fernandes, T.F.; Forte, J.; Karakassis, I.; Muxika, I.; Nickell, T.D.; et al. Assessing the suitability of a range of benthic indices in the evaluation of environmental impact of fin and shellfish aquaculture located in sites across Europe. Aquaculture 2009, 293, 231–240. [Google Scholar] [CrossRef] [Green Version]
- Read, P.; Fernandes, T. Management of environmental impacts of marine aquaculture in Europe. Aquaculture 2003, 226, 139–163. [Google Scholar] [CrossRef]
- Lester, S.E.; Stevens, J.M.; Gentry, R.R.; Kappel, C.V.; Bell, T.W.; Costello, C.J.; Gaines, S.D.; Kiefer, D.A.; Maue, C.C.; Rensel, J.E.; et al. Marine spatial planning makes room for offshore aquaculture in crowded coastal waters. Nat. Commun. 2018, 9, 945. [Google Scholar] [CrossRef] [PubMed]
- Sarà, G.; Mangano, M.C.; Johnson, M.; Mazzola, A. Integrating multiple stressors in aquaculture to build the blue growth in a changing sea. Hydrobiologia 2018, 809, 5–17. [Google Scholar] [CrossRef]
- Van den Burg, S.W.K.; Kamermans, P.; Blanch, M.; Pletsas, D.; Poelman, M.; Soma, K.; Dalton, G. Business case for mussel aquaculture in offshore wind farms in the North Sea. Mar. Policy 2017. [Google Scholar] [CrossRef]
- Jansen, H.M.; van den Burg, S.; Bolman, B.; Jak, R.G.; Kamermans, P.; Poelman, M.; Stuiver, M. The feasibility of offshore aquaculture and its potential for multi-use in the North Sea. Aquacult. Int. 2016. [Google Scholar] [CrossRef]
- Gentry, R.R.; Froehlich, H.E.; Grimm, D.; Kareiva, P.; Parke, M.; Rust, M.; Gaines, S.D.; Halpern, B.S. Mapping the global potential for marine aquaculture. Nat. Ecol. Evol. 2017, 1, 1317–1324. [Google Scholar] [CrossRef] [PubMed]
- Pirlet, H.; Claus, S.; Copejans, E.D.C.E.; Guanche García, R.; Møhlenberg, F.; Rappé, K.; Schouten, J.-J.; Zanuttigh, B. The Mermaid Project—Innovative Multi-Purpose Offshore Platforms; Flanders Marine Institute (VLIZ): Ostend, Belgium, 2014; p. 20. [Google Scholar]
- Quevedo, E.; Delory, E.; Castro, A.; Llinás, O.; Hernández, J. Modular multi-purpose offshore platforms, the TROPOS project approach. In Proceedings of the 4th International Conference on Ocean Energy, Dublin, Ireland, 17–19 October 2012. [Google Scholar]
- Olsen, O.T. The Piscatorial Atlas of the North Sea, English and St. George’s Channels; Grimsby: London, UK, 1883. [Google Scholar]
- Gercken, J.; Schmidt, A. Current Status of the European Oyster (Ostrea edulis) and Possibilities for Restoration in the German North Sea 2014; BFN Report; Bundesamt für Naturschutz: Bonn, Germany, 2014. [Google Scholar]
- Houziaux, J.-S.; Kerckhof, F.; Degrendele, K.; Roche, M.; Norro, A. The Hinder Banks Yet an Important Region for the Belgian Marine Biodiversity? Final Report HINDERS Project; Belgian Science Policy Office: Brussels, Belgium, 2008; p. 249. Available online: http://www.belspo.be/belspo/home/publ/pub_ostc/EV/rappEV45_en.pdfS (accessed on 28 August 2018).
- Bruno, J.F.; Stachowicz, J.J.; Bertness, M.D. Inclusion of facilitation into ecological theory. Trends Ecol. Evol. 2003, 18, 119–125. [Google Scholar] [CrossRef]
- Bouma, T.J.; Olenin, S.; Reise, K.; Ysebaert, T. Ecosystem engineering and biodiversity in coastal sediments: Posing hypotheses. Helgol. Mar. Res. 2009, 63, 95–106. [Google Scholar] [CrossRef]
- Lenihan, H.S.; Peterson, C.H. How habitat degradation through fishery disturbance enhances impacts of hypoxia on oyster reefs. Ecol. Appl. 1998, 8, 128–140. [Google Scholar] [CrossRef]
- zu Ermgassen, P.S.E.; Grabowski, J.H.; Gair, J.R.; Powers, S.P. Quantifying fish and mobile invertebrate production from a threatened nursery habitat. J. Appl. Ecol. 2016, 53, 596–606. [Google Scholar] [CrossRef]
- Baggett, L.P.; Powers, S.P.; Brumbaugh, R.; Coen, L.D.; DeAngelis, B.; Greene, J.; Hancock, B.; Morlock, S. Oyster Habitat Restoration Monitoring and Assessment Handbook; The Nature Conservancy: Arlington, VA, USA, 2014; 96p, Available online: http://www.oyster-restoration.org/wp-content/uploads/2014/01/Oyster-Habitat-Restoration-Monitoring-and-Assessment-Handbook.pdf (accessed on 28 August 2018).
- Zwerschke, N.; Emmerson, M.C.; Roberts, D.; O’Connor, N.E. Benthic assemblages associated with native and non-native oysters are similar. Mar. Pollut. Bull. 2016, 111, 305–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smyth, D.; Roberts, D. The European oyster (Ostrea edulis) and its epibiotic succession. Hydrobiologia 2010, 655, 25–36. [Google Scholar] [CrossRef]
- Newell, R.I.E. Ecosystem influences of natural and cultivated populations of suspension-feeding bivalve molluscs: A review. J. Shellfish Res. 2004, 23, 51–61. [Google Scholar]
- Kellogg, M.L.; Cornwell, J.C.; Owens, M.S.; Paynter, K.T. Denitrification and nutrient assimilation on a restored oyster reef. Mar. Ecol. Prog. Ser. 2013, 480, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Dame, R.F.; Prins, T. Bivalve carrying capacity in coastal ecosystems. Aquat. Ecol. 1997, 31, 409–421. [Google Scholar] [CrossRef]
- Van der Zee, E.; Heide, T.; Donadi, S.; Eklöf, J.; Eriksson, B.; Olff, H.; Veer, H.; Piersma, T. Spatially extended habitat modification by intertidal reef-building bivalves has implications for consumer-resource interactions. Ecosystems 2012, 15, 664–673. [Google Scholar] [CrossRef]
- Van Oevelen, D.; Duineveld, G.; Lavaleye, M.; Mienis, F.; Soetaert, K.; Heip, C.H.R. The cold-water coral community as a hot spot for carbon cycling on continental margins: A food-web analysis from Rockall Bank (northeast Atlantic). Limnol. Oceanogr. 2009, 54, 1829–1844. [Google Scholar] [CrossRef]
- De Smet, B.; van Oevelen, D.; Vincx, M.; Vanaverbeke, J.; Soetaert, K. Lanice conchilega structures carbon flows in soft-bottom intertidal areas. Mar. Ecol. Prog. Ser. 2016, 552, 47–60. [Google Scholar] [CrossRef] [Green Version]
- Van der Heide, T.; Tielens, E.; van der Zee, E.M.; Weerman, E.J.; Holthuijsen, S.; Eriksson, B.K.; Piersma, T.; van de Koppel, J.; Olff, H. Predation and habitat modification synergistically interact to control bivalve recruitment on intertidal mudflats. Boil. Conserv. 2014, 172, 163–169. [Google Scholar] [CrossRef] [Green Version]
- Ysebaert, T.; Hart, M.; Herman, P.M.J. Impacts of bottom and suspended cultures of mussels Mytilus spp. on the surrounding sedimentary environment and macrobenthic biodiversity. Helgol. Mar. Res. 2009, 63, 59–74. [Google Scholar] [CrossRef]
- Puls, W.; Sündermann, J. Simulation of Suspended Sediment Dispersion in the North Sea. In Residual Currents and Long-Term Transport; Coastal and Estuarine Studies; Springer: Berlin, Germany, 1990; Volume 38. [Google Scholar]
- Korringa, P. The shells of Ostrea edulis as a habitat. Arch. Neerl. Zoöl. 1954, 10, 32–146. [Google Scholar] [CrossRef]
- Reise, K.; Beusekom, J.E.E. Interactive effects of global and regional change on a coastal ecosystem. Helgol. Mar. Res. 2008, 62, 85–91. [Google Scholar] [CrossRef]
- Ministry of Economic Affairs. 2014 Nature Ambition Large Water Bodies. Available online: www.minez.nl (accessed on 28 August 2018).
- Ministry of Economic Affairs. 2013 Implementation Agenda Natural Capital. Available online: www.minez.nl (accessed on 28 August 2018).
- High Level Panel of Experts. Sustainable Fisheries and Aquaculture for Food Security and Nutrition: A Report by the High Level Panel of Experts on Food Security and Nutrition of the Committee on World Food Security; FAO: Rome, Italy, 2014. [Google Scholar]
- Golden, C.D.; Allison, E.H.; Cheung, W.W.L.; Dey, M.M.; Halpern, B.S.; McCauley, D.J. Nutrition: Fall in fish catch threatens human health. Nature 2016, 534, 317–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allison, E.H.; Béné, C.; Andrew, N.L. Poverty reduction as a means to enhance resilience in small-scale fisheries. In Small-Scale Fisheries Management—Frameworks and Approaches for the Developing World; Pomeroy, R.S., Andrew, N.L., Eds.; CABI: Wallingford, UK, 2011; pp. 216–238. [Google Scholar]
- Béné, C.; Arthur, R.; Norbury, H.; Allison, E.H.; Beveridge, M.; Bush, S.; Campling, L.; Leschen, W.; Little, D.; Squires, D.; et al. Contribution of fisheries and aquaculture to food security and poverty reduction: Assessing the current evidence. World Dev. 2016, 79, 177–196. [Google Scholar] [CrossRef]
- Beveridge, M.C.M.; Thilsted, S.H.; Phillips, M.J.; Metian, M.; Troell, M.; Hall, S.J. Meeting the food and nutrition needs of the poor: The role of fish and the opportunities and challenges emerging from the rise of aquaculture. J. Fish Boil. 2013, 83, 1067–1084. [Google Scholar] [CrossRef] [PubMed]
- Kolding, J.; van Zwieten, P.; Marttin, F.; Poulain, F. Fisheries in the Drylands of Sub-Sahara. Fish Come with the Rains; Building Resilience for Fisheries Dependant Livelihoods to Enhance Food Security and Nutrition in the Drylands; FAO Report FIAF/FIAP/C1118(En).7; Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 2016. [Google Scholar]
- Bogard, J.R.; Thilsted, S.H.; Marks, G.C.; Abdul Wahab, M.; Hossain, M.A.R.; Jakobsen, J.; Stangoulis, J. Nutrient composition of important fish species in Bangladesh and potential contribution to recommended nutrient intakes. J. Food Compos. Anal. 2015, 42, 120–133. [Google Scholar] [CrossRef] [Green Version]
- Kurien, J. Responsible Fish Trade and Food Security; FAO Fisheries Technical Paper 456; Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 2005. [Google Scholar]
- Pelletier, N.; Audsley, E.; Brodt, S.; Garnett, T.; Henriksson, P.; Kendall, A.; Kramer, K.J.; Murphy, D.; Nemecek, T.; Troell, M. Energy Intensity of Agriculture and Food Systems. Annu. Rev. Environ. Resour. 2011, 36, 223–246. [Google Scholar] [CrossRef]
- Hilborn, R.; Tellier, P. The Environmental Cost of N. Z. Food Production; New Zealand Seafood Industry Council Ltd.: Wellington, New Zealand, 2012; ISBN 978-0473-17824-6. [Google Scholar]
- Hall, S.J.; Delaporte, A.; Phillips, M.J.; Beveridge, M.; O’Keefe, M. Blue Frontiers: Managing the Environmental Costs of Aquaculture; The WorldFish Center: Penang, Malaysia, 2011. [Google Scholar]
- Garcia, S.M.; Kolding, J.; Rice, J.; Rochet, M.J.; Zhou, S.; Arimoto, T.; Beyer, J.E.; Borges, L.; Bundy, A.; Dunn, D. Reconsidering the consequences of selective fisheries. Science 2012, 335, 1045–1047. [Google Scholar] [CrossRef] [PubMed]
- Smaal, A.C.; Kamermans, P.; van der Have, T.M.; Engelsma, M.; Sas, H.W.J. Feasibility of Flat Oyster (Ostrea edulis) Restoration in the Dutch Part of the North Sea; IMARES Report C028/15; IMARES Wageningen UR: Ijmuiden, The Netherlands, 2015. [Google Scholar]
- Smaal, A.; Kamermans, P.; Kleissen, F.; van Duren, L.; van der Have, T. Flat Oysters on Offshore Wind Farms. Opportunities for the Development of flat Oyster Populations on Existing and Planned Wind Farms in the Dutch Section of the North Sea; Wageningen Marine Research Rapport C052/17; Wageningen Marine Research: Yerseke, The Netherlands, 2017. [Google Scholar]
- Kerckhof, F.; Coolen, J.W.P.; Rumes, B.; Degraer, S. Recent findings of wild European flat oysters Ostrea edulis (Linnaeus, 1758) in Belgian and Dutch offshore waters: New perspectives for offshore oyster reef restoration in the southern North Sea Belgian. J. Zoöl. 2018. [Google Scholar] [CrossRef]
- Burke, K.; Bataller, É.; Miron, G. Spat collection of a non-native bivalve species (European oyster, Ostrea edulis) off the Eastern Canadian coast. J. Shellfish Res. 2008, 27, 345–353. [Google Scholar] [CrossRef]
- Smyth, D.; Mahon, A.M.; Roberts, D.; Kregting, L. Settlement of Ostrea edulis is determined by the availability of hard substrata rather than by its nature: Implications for stock recovery and restoration of the European oyster. Aquat. Conserv. Mar. Freshw. Ecosyst. 2018. [Google Scholar] [CrossRef]
- Walne, P.R. Culture of Bivalve moLluscs: 50 Years of Experience at Conwy; Fishing News Books Ltd.: Farnham, UK, 1974; 189p. [Google Scholar]
- O’Foighil, D. Role of spermatozeugmata in the spawning ecology of the brooding oyster Ostrea edulis. Gamete Res. 1989, 24, 219–228. [Google Scholar] [CrossRef] [PubMed]
- Muus, B.J.; Dahlstrom, P. Meeresfische; BLV: Munchen, Germany, 1973. [Google Scholar]
- Jackson, A. Ostrea edulis. Native oyster. Marine Life Information Network: Biology and Sensitivity Key Information Sub-Programmem [on-line]; Marine Biological Association: Plymouth, UK, 2007. [Google Scholar]
- Berghahn, R.; Ruth, M. The disappearance of oysters from the Wadden Sea: A cautionary tale for no-take zones. Aquat. Conserv. Mar. Freshw. Ecosyst. 2005, 15, 91–104. [Google Scholar] [CrossRef]
- Jordan-Cooley, W.C.; Lipcius, R.N.; Shaw, L.B.; Shen, J.; Shi, J. Bistability in a differential equation model of oyster population and sediment volume. J. Theor. Boil. 2011, 289, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, R.J.; Roberts, D. Commercial oyster stocks as a potential source of larvae in the regeneration of Ostrea edulis in Strangford Lough, Northern Ireland. JMBA 2006, 86, 153–159. [Google Scholar] [CrossRef]
- Vera, M.; Carlsson, J.; Carlsson, J.E.L.; Cross, T.; Lynch, S.; Kamermans, P.; Villalba, A.; Culloty, S.; Martinez, P. Current genetic status, temporal stability and structure of the remnant wild European flat oyster populations: Conservation and restoring implications. Mar. Biol. 2016, 163, 239. [Google Scholar] [CrossRef]
- Smyth, D.; Kregting, L.; Elsäßer, B.; Kennedy, R.; Roberts, D. Using particle dispersal models to assist in the conservation and recovery of the overexploited native oyster (Ostrea edulis) in an enclosed sea lough. JSR 2016, 108, 50–59. [Google Scholar] [CrossRef]
- Capelle, J.J. Restoration of the Oyster Ostrea edulis L. in the Wadden Sea; Wageningen IMARES, Rapport; IMARES: Yerseke, The Netherlands, 2008; p. 61. [Google Scholar]
- Haure, J.; Penisson, C.; Bougrier, S.; Baud, J.P. Influence of temperature on clearance and oxygen consumption rates of the flat oyster Ostrea edulis: Determination of allometric coefficients. Aquaculture 1998, 169, 211–224. [Google Scholar] [CrossRef]
- Newell, R.C.; Johnson, L.G.; Kofoed, L.H. Adjustment of the components of energy balance in response to temperature change in Ostrea edulis. Oecologia 1977, 30, 97–110. [Google Scholar] [CrossRef] [PubMed]
- Lubet, P. Ecophysiologie de la reproduction chez les mollusques lamellibranches. Haliotis 1976, 7, 49–55. [Google Scholar]
- Buxton, C.D.; Newell, R.C.; Field, J.G. Response-Surface Analysis of the Combined Effects of Exposure and Acclimation Temperatures on Filtration, Oxygen Consumption and Scope for Growth in the Oyster. Mar. Ecol. Prog. Ser. 1981, 6, 73–82. [Google Scholar] [CrossRef]
- Davis, H.C.; Calabrese, A. Survival and growth of larvae of the European oyster (Ostrea edulis L.) at different temperatures. Boil. Bull. 1969, 136, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Hoek, P.P.C. Rapport Over de Oorzaken van den Achteruitgang in Hoedanigheid van de Zeeuwsche oester; Ministerie van Waterstaat, Handel en Nijverheid: ‘s-Gravenhage, The Netherlans, 1902.
- Child, A.R.; Laing, I. Comparative low temperature tolerance of small juvenile European, Ostrea edulis L.; Pacific oysters, Crassostrea gigas Thunberg. Aquac. Res. 1998, 29, 103–113. [Google Scholar] [CrossRef]
- Davis, H.C.; Ansell, A.D. Survival and growth of larvae of the European oyster (Ostrea edulis L.) at lowered salinities. Boil. Bull. 1962, 122, 33–39. [Google Scholar] [CrossRef]
- Hutchinson, S.; Hawkins, L.E. Quantification of the physiological responses of the European flat oyster, Ostrea edulis L. to temperature and salinity. J. Molluscan Stud. 1992, 58, 215–226. [Google Scholar] [CrossRef]
- Hayward, P.J.; Ryland, J.S. Handbook of the Marine Fauna of North-West Europe; Oxford University Press: New York, NY, USA, 1998. [Google Scholar]
- Drinkwaard, A.C. Current velocity as an ecological factor in shell growth of Ostrea edulis. ICES Shellfish Committee 1961, 47, 1–3. [Google Scholar]
- Grant, J.; Enright, C.T.; Griswold, A. Resuspension and growth of Ostrea edulis: A field experiment. Mar. Boil. 1990, 104, 51–59. [Google Scholar] [CrossRef]
- Tillin, H.M.; Hull, S.C. Tools for Appropriate Assessment of Fishing and Aquaculture Activities in Marine and Coastal Natura 2000 Sites; Report VII: Intertidal and Subtidal Reefs, Report No. R. 2074; ABPmer for the Marine Institute: Galway, Ireland, 2013. [Google Scholar]
- Hawkins, A.J.S.; Bayne, B.L.; Bougrier, S.; Héral, M.; Iglesias, J.I.P.; Navarro, E.; Smith, R.F.M.; Urrutia, M.B. Some general relationships in comparing the feeding physiology of suspensionfeeding bivalve molluscs. J. Exp. Mar. Boil. Ecol. 1998, 219, 87–103. [Google Scholar] [CrossRef]
- Barillé, L.; Prou, J.; Heral, M.; Razet, D. Effects of high natural seston concentrations on the feeding, selection, and absorption of the oyster Crassostrea gigas Thunberg. JEMBE 1997, 2012, 149–172. [Google Scholar] [CrossRef]
- Millican, P.F.; Helm, M.M. Effects of nutrition on larvae production in the European flat Oyster, Ostrea edulis. Aquaculture 1994, 123, 83–94. [Google Scholar] [CrossRef]
- Davis, J.C. Minimal Dissolved Oxygen Requirements of Aquatic Life with Emphasis on Canadian Species: A Review. J. Fish. Board Can. 1975, 32, 2295–2332. [Google Scholar] [CrossRef]
- Borsje, B.W.; Bouma, T.J.; Rabaut, M.; Herman, P.M.J.; Hulscher, S.J.M.H. Formation and erosion of biogeomorphological structures: A model study on the tube-building polychaete Lanice conchilega. Limnol. Oceanogr. 2014, 59, 1297–1309. [Google Scholar] [CrossRef]
- Buijsman, M.C.; Ridderinkhof, H. Long-term evolution of sand waves in the Marsdiep inlet. II: Relation to hydrodynamics. Cont. Shelf Res. 2008, 28, 1202–1215. [Google Scholar] [CrossRef]
- Dorst, L.L.; Roos, P.C.; Hulscher, S.J.M.H.; Lindenbergh, R.C. The estimation of sea floor dynamics from bathymetric surveys of a sand wave area. J. Appl. Geodesy 2009, 3, 97–120. [Google Scholar] [CrossRef]
- Abelson, A.; Weihs, D.; Loya, Y. Hydrodynamic impediments to settlement of marine propagules, and adhesive-filament solutions. Limnol. Oceanogr. 1994, 39, 164–169. [Google Scholar] [CrossRef]
- Crimaldi, J.P.; Thompson, J.K.; Rosman, J.H.; Lowe, R.J.; Koseff, J.R. Hydrodynamics of larval settlement: The influence of turbulent stress events at potential recruitment sites. Limnol. Oceanogr. 2002, 47, 1137–1151. [Google Scholar] [CrossRef] [Green Version]
- Hendriks, I.E.; van Duren, L.A.; Herman, P.M.J. Turbulence levels in a flume compared to the field. J. Sea Res. 2006, 55, 15–29. [Google Scholar] [CrossRef]
- Grabowski, R.C.; Droppo, I.G.; Wharton, G. Erodibility of cohesive sediment: The importance of sediment properties. Earth-Sci. Rev. 2011, 105, 101–120. [Google Scholar] [CrossRef]
- Wright, L.D.; Schaffner, L.C.; Maa, J.P.-Y. Biological mediation of bottom boundary layer processes and sediment suspension in the lower Chesapeake Bay. Mar. Geol. 1997, 141, 27–50. [Google Scholar] [CrossRef]
- McCave, I.N. Sand waves in the North Sea off the coast of Holland. Mar. Geol. 1971, 10, 199–225. [Google Scholar] [CrossRef]
- Van Dijk, T.A.G.P.; Kleinhans, M.G. Processes controlling the dynamics of compound sand waves in the North Sea, Netherlands. J. Geophys. Res. Earth Surf. 2005, 110. [Google Scholar] [CrossRef] [Green Version]
- Damen, J.M.; van Dijk, T.A.G.P.; Hulscher, S.J.M.H. Spatially Varying Environmental Properties Controlling Observed Sand Wave Morphology. J. Geophys. Res. 2018, 123, 19. [Google Scholar] [CrossRef]
- ICONA. Noordzeeatlas Voor Het Nederlandse Beleid en Beheer; ICONA Den Haag; Stadsuitgeverij: Amsterdam, The Netherlands, 1992. [Google Scholar]
- Gerritsen, H.; Vos, R.J.; Van Der Kaaij, T.; Lane, A.; Boon, J.G. Suspended sediment modelling in a shelf sea (North Sea). Coast. Eng. 2000, 41, 317–352. [Google Scholar] [CrossRef]
- Van der Kaaij, T.; van Kessel, T.; Troost, T.; van Duren, L.A.; Villars, M.T. Modelondersteuning MER Winning Suppletie- en Ophoogzand Noordzee 2018–2027; Modelvalidatie 1230888-002; Deltares: Delft, The Netherlands, 2017. [Google Scholar]
- Laing, I.; Walker, P.; Areal, F. A Feasibility Study of Native Oyster (Ostrea edulis) Stock Regeneration in the United Kingdom; CARD Project Report FC1016; Centre for Environment, Fisheries & Aquaculture Science: Lowestoft, UK, 2005.
- Sawusdee, A.; Jensen, A.C.; Collins, K.J.; Hauton, C. Improvements in the physiological performance of European flat oysters Ostrea edulis (Linnaeus, 1758) cultured on elevated reef structures: Implications for oyster restoration. Aquaculture 2015, 444, 41–48. [Google Scholar] [CrossRef]
- Snelgrove, P.V.R.; Butman, C.A. Animal-sediment relationships revisited: Cause versus effect. Oceanogr. Mar. Boil. Annu. Rev. 1994, 32, 111–177. [Google Scholar]
- Houziaux, J.-S.; Fettweis, M.; Francken, F.; van Lancker, V. Historic (1900) seafloor composition in the Belgian–Dutch part of the North Sea: A reconstruction based on calibrated visual sediment descriptions. Cont. Shelf Res. 2011, 31, 1043–1056. [Google Scholar] [CrossRef]
- Wasson, K. Informing Olympia Oyster Restoration: Evaluation of Factors That Limit Populations in a California Estuary. Wetlands 2010, 30, 449–459. [Google Scholar] [CrossRef]
- Peirano, A. Wrecks on the Bottom: Useful, Ecological Sentinels? Mar. Technol. Soc. J. 2013, 47, 118–127. [Google Scholar] [CrossRef]
- Petersen, J.K.; Maar, M.; Ysebaert, T.; Herman, P.M.J. Near-bed gradients in particles and nutrients above a mussel bed in the Limfjorden: Influence of physical mixing and mussel filtration. Mar. Ecol. Prog. Ser. 2013, 490, 137–146. [Google Scholar] [CrossRef]
- Wiles, P.J.; van Duren, L.A.; Haese, C.; Larsen, J.; Simpson, J.H. Stratification and mixing in Limfjorden in relation to mussel culture. J. Mar. Syst. 2006, 60, 129–134. [Google Scholar] [CrossRef]
- Van Leeuwen, S.; Tett, P.; Mills, D.; van der Molen, J. Stratified and nonstratified areas in the North Sea: Long-term variability and biological and policy implications. J. Geophys. Res. 2015, 120, 4670–4686. [Google Scholar] [CrossRef] [Green Version]
- Greenwood, N.; Parker, E.R.; Fernand, L.; Sivyer, D.B.; Weston, K.; Painting, S.J.; Kröger, S.; Forster, R.M.; Lees, H.E.; Mills, D.K.; et al. Detection of low bottom water oxygen concentrations in the North Sea; implications for monitoring and assessment of ecosystem health. Biogeosciences 2010, 7, 1357–1373. [Google Scholar] [CrossRef] [Green Version]
- Queste, B.Y.; Fernand, L.; Jickells, T.D.; Heywood, K.J.; Hind, A.J. Drivers of summer oxygen depletion in the central North Sea. Biogeosciences 2016, 13, 1209–1222. [Google Scholar] [CrossRef] [Green Version]
- Cronin, K.; Plieger, R.; Gaytan Aguilar, S.; de Lima Rego, J.R.; Blaas, M. MoS2-II: Deterministic Model Calibration: Updates of the ZUNO-DD Hydrodynamic and SPM Model Deltares Report; Rep. No. 1204561; Deltares: Delft, The Netherlands, 2013. [Google Scholar]
- Korringa, P. Relations between the Moon and Periodicity in the Breeding of Marine Animals. Ecol. Monogr. 1947, 17, 347–381. [Google Scholar] [CrossRef]
- Sas, H.; Kamermans, P.; van der Have, T.; Lengkeek, W.; Smaal, A. Shellfish Reef Restoration Pilots, Voordelta, The Netherlands; Annual Report; ARK Natuurontwikkeling: Nijmegen, The Netherlands, 2016. [Google Scholar]
- Sas, H.; Kamermans, P.; van der Have, T.; Christianen, M. Shellfish reef Restoration Pilots, Voordelta, The Netherlands; Annual Report; ARK Natuurontwikkeling: Nijmegen, The Netherlands, 2017. [Google Scholar]
- Christianen, M.J.A.; Bergsma, J.H.; Coolen, J.W.P.; Didderen, K.; Dorenbosch, M.; Driessen, F.M.F.; Kamermans, P.; Reuchlin-Hugenholtz, E.; Sas, H.; Smaal, A.; et al. Return of the native facilitated by the invasive? Population composition, substrate preferences and epibenthic species richness of a recently discovered shellfish reef with native European flat oysters (Ostrea edulis) in the North Sea. Mar. Boil. Res. 2018, 14, 590–597. [Google Scholar] [CrossRef]
- Petersen, T.U.; Mutlu Sumer, B.; Fredsøe, J.; Raaijmakers, T.C.; Schouten, J.J. Edge scour at scour protections around piles in the marine environment—Laboratory and field investigation. Coast. Eng. 2015, 106, 42–72. [Google Scholar] [CrossRef]
- Pogoda, B.; Buck, B.H.; Hagen, W. Growth performance and condition of oysters (Crassostrea gigas and Ostrea edulis) farmed in an offshore environment (North Sea, Germany). Aquaculture 2011, 319, 484–492. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Survival | Growth | Reproduction | Recruitment | |
|---|---|---|---|---|
| Sea bed shear stress | – | |||
| Sea bed motion | – | |||
| Concentration of suspended particles | – | |||
| Larval retention | + | |||
| Coarse sediment | + |
| Environmental Factor | Response Variable | Range | Optimum | Reference |
|---|---|---|---|---|
| Temperature (°C) | Survival adults | <30 | [66] | |
| Growth adults | 6–19 | 7–14 | [66,67,68] | |
| 17 | [69] | |||
| Gonad development | 7–14 | [68] | ||
| Survival and growth larvae | 10–31 | 25–27 | [70] | |
| Spat fall | >18.5 | [71] | ||
| Survival spat | >3 | [72] | ||
| Salinity (PSU) | Survival and growth larvae | 20–39.5 | 25–35 | [73] |
| Growth adults | >19 | [74] | ||
| Water depth (m) | Survival adults | <–80 | [75] | |
| Current velocity (m/s) | Survival adults | <0.25 | 0.03 | [76] |
| Sea bed motion (cm/day) | Survival spat and adults | <0.8 | [77] | |
| Sediment composition | Survival adults | Firm silty sand or silty gravel with shells and stones | [78] | |
| Suspended matter concentration (mg/L) | Food intake | <90 | [79,80] | |
| Food supply (chla ug/L) | Gonad development | 1.68 | [81] | |
| Oxygen concentration (mg/L) | Survival adults | >0.5 | [82] |
| Wind Farm | Sea Bed Shear Stress | Suspended | Matter | Sediment | Historical Occurrence | Larval Retention | Sum |
|---|---|---|---|---|---|---|---|
| avg | max | avg | |||||
| Borssele | 1 | 2 | 2 | 1 | 2 | 2 | 10 |
| HK-Zuid | 1 | 2 | 2 | 1 | 1 | 1 | 8 |
| OWEZ | 1 | 1 | 2 | 1 | 1 | 2 | 8 |
| HK-Noord | 1 | 2 | 2 | 1 | 1 | 1 | 8 |
| Luchterduinen | 1 | 2 | 2 | 1 | 1 | 2 | 9 |
| Princes Amalia | 1 | 2 | 2 | 1 | 1 | 1 | 8 |
| Buitengaats | 2 | 2 | 2 | 1 | 2 | 1 | 10 |
| Zee-energie | 2 | 2 | 2 | 2 | 2 | 2 | 12 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kamermans, P.; Walles, B.; Kraan, M.; Van Duren, L.A.; Kleissen, F.; Van der Have, T.M.; Smaal, A.C.; Poelman, M. Offshore Wind Farms as Potential Locations for Flat Oyster (Ostrea edulis) Restoration in the Dutch North Sea. Sustainability 2018, 10, 3942. https://doi.org/10.3390/su10113942
Kamermans P, Walles B, Kraan M, Van Duren LA, Kleissen F, Van der Have TM, Smaal AC, Poelman M. Offshore Wind Farms as Potential Locations for Flat Oyster (Ostrea edulis) Restoration in the Dutch North Sea. Sustainability. 2018; 10(11):3942. https://doi.org/10.3390/su10113942
Chicago/Turabian StyleKamermans, Pauline, Brenda Walles, Marloes Kraan, Luca A. Van Duren, Frank Kleissen, Tom M. Van der Have, Aad C. Smaal, and Marnix Poelman. 2018. "Offshore Wind Farms as Potential Locations for Flat Oyster (Ostrea edulis) Restoration in the Dutch North Sea" Sustainability 10, no. 11: 3942. https://doi.org/10.3390/su10113942