Overwintered Drosophila suzukii Are the Main Source for Infestations of the First Fruit Crops of the Season

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Identification and Natural Infestation of Early Spring Hosts
2.2. Natural Infestation of A. japonica by D. suzukii over Time and Phenotype of Emerged Adults
2.3. Performance of D. suzukii on Early Spring Hosts in Controlled Conditions
2.4. Field Monitoring Program
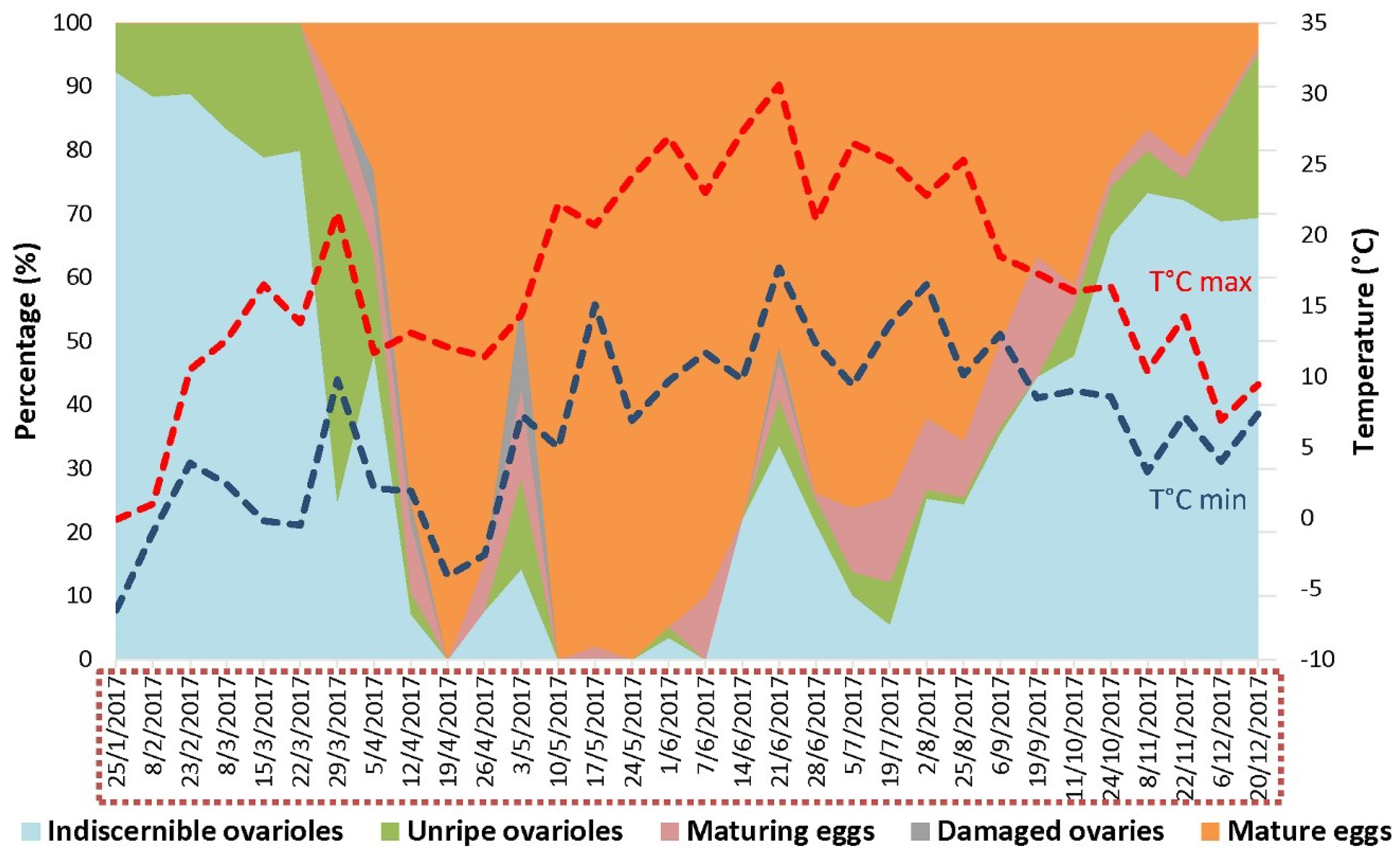
2.4.1. Ovary Dissection
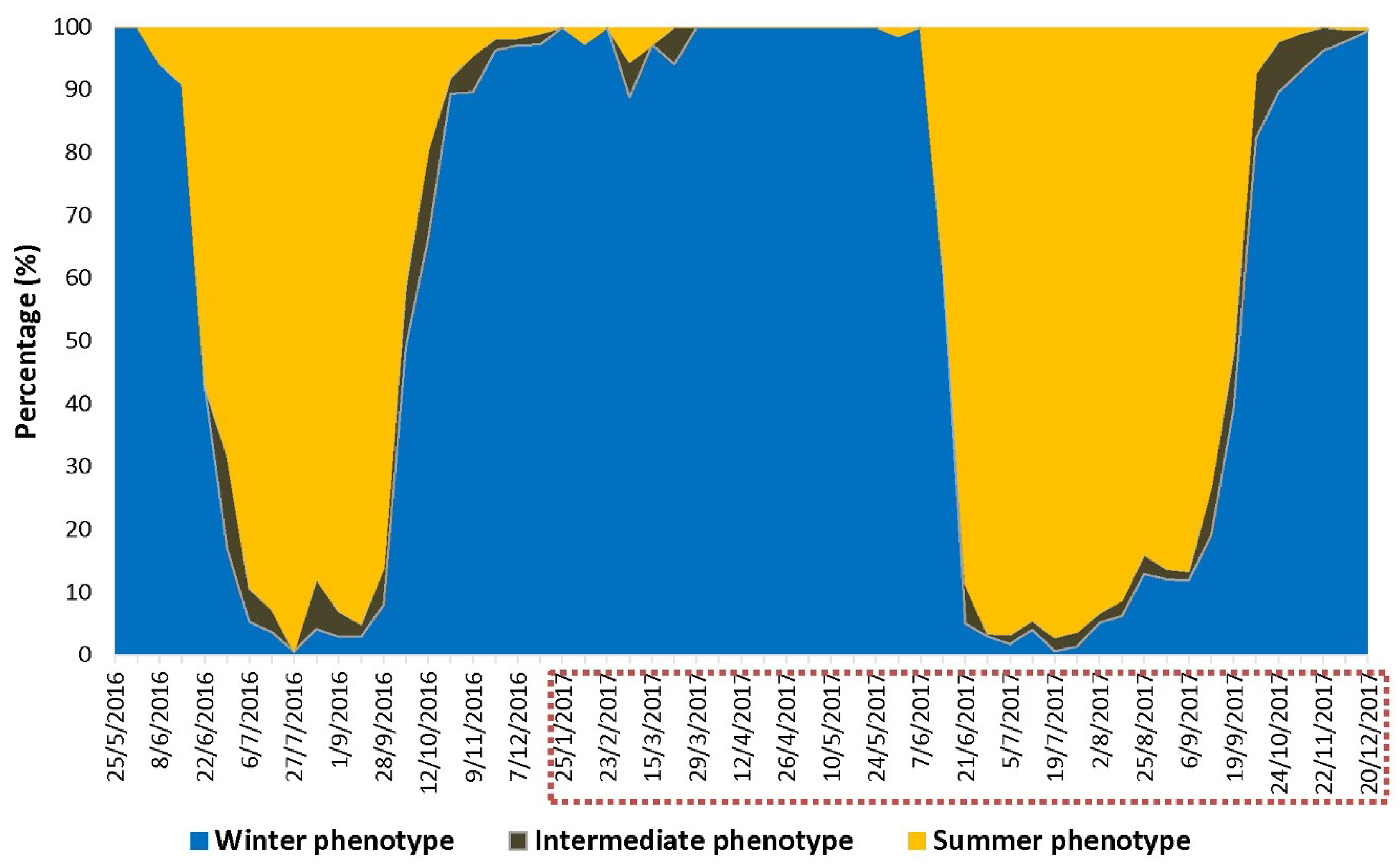
2.4.2. Phenotype Assessment
2.5. Temperature Model
2.6. Data Analysis
3. Results
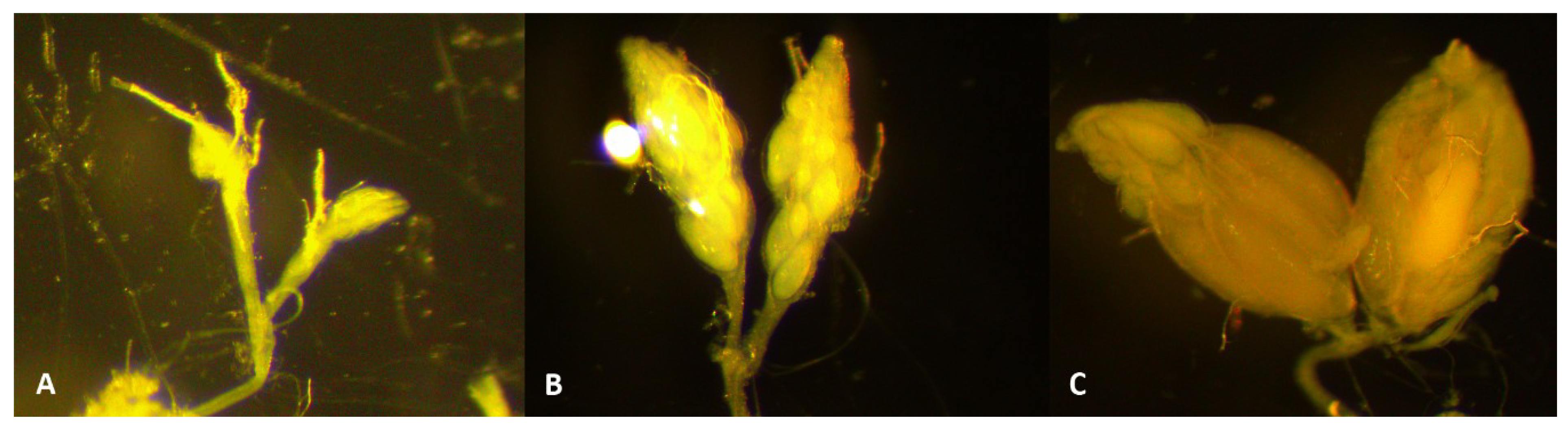
3.1. A. japonica Is Highly Infested by D. suzukii in Nature, but with a Low and Variable Egg-to-Adult Survival
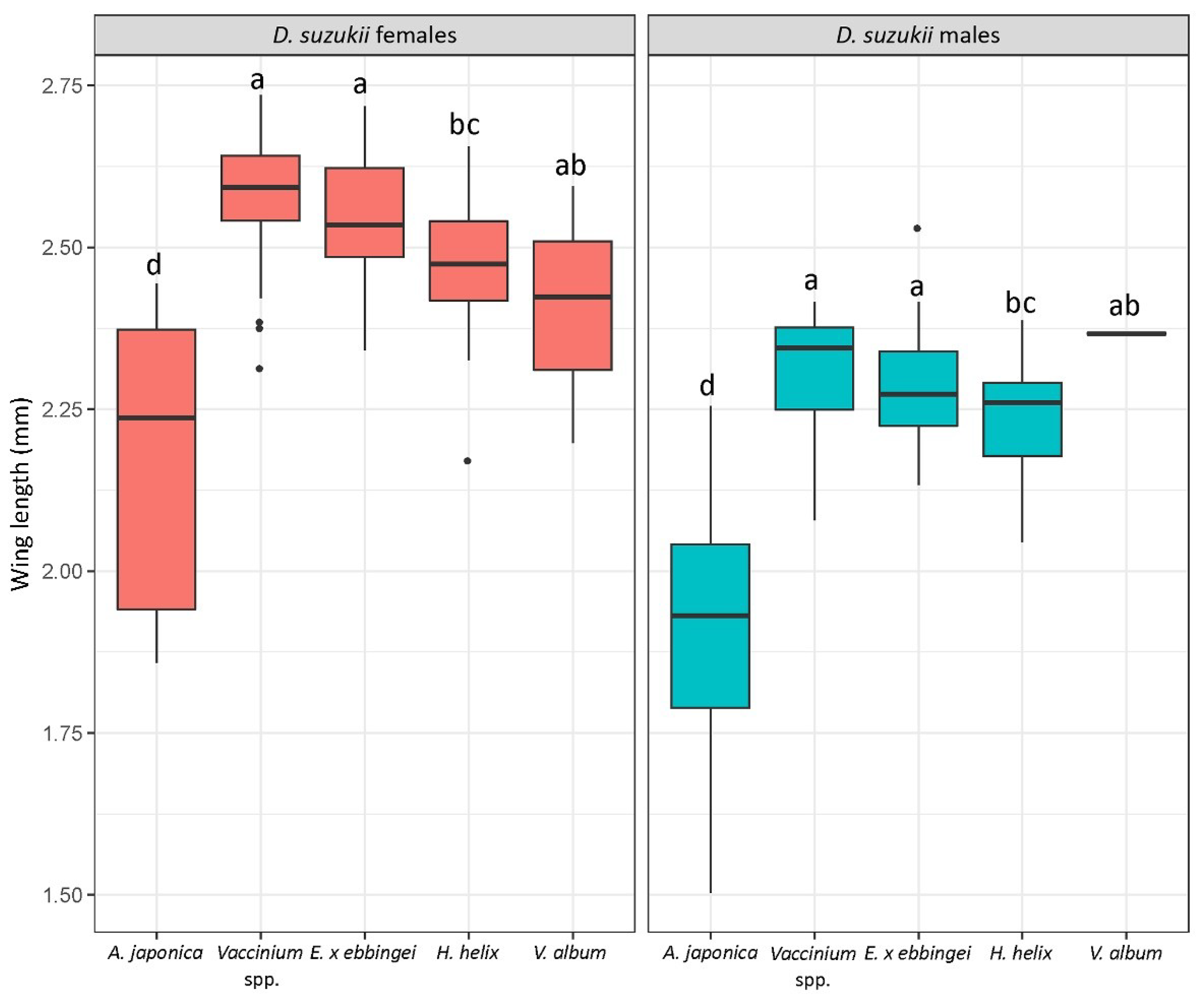
3.2. D. suzukii Infest A. japonica from Early to Late Spring and a Few Summer-Morph Flies Emerge from This Host
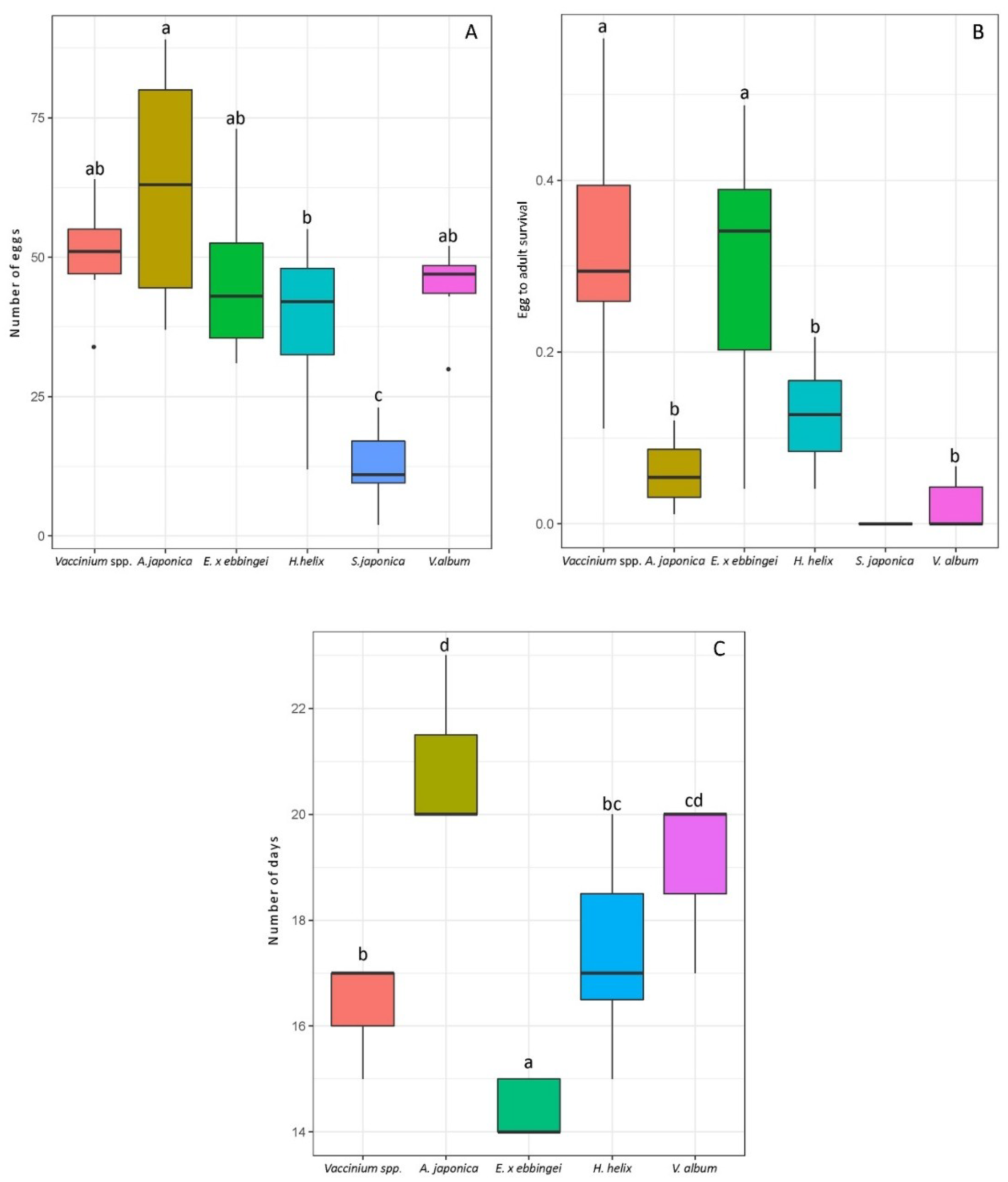
3.3. Laboratory Assays Confirm the Field Observations
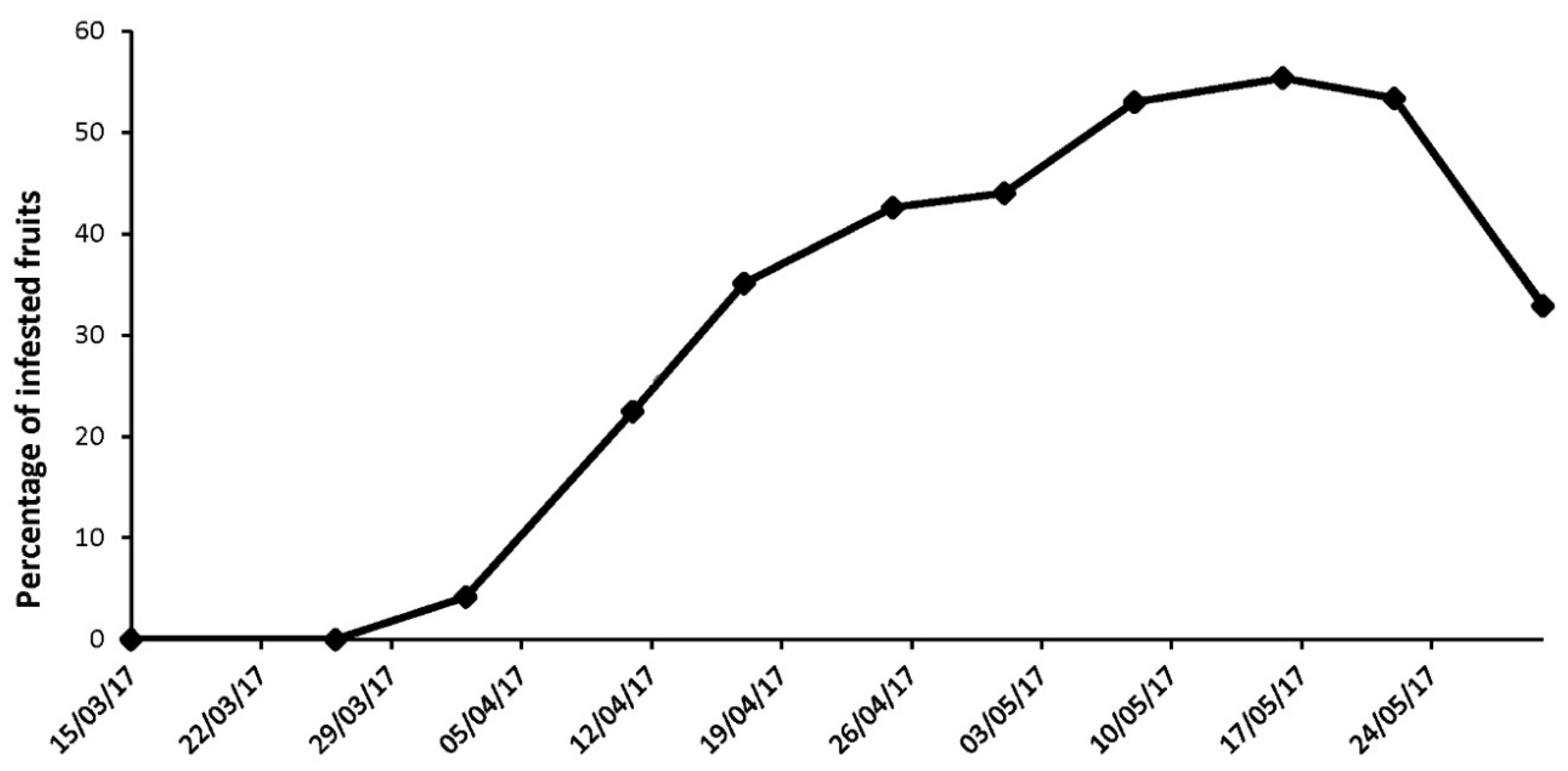
3.4. Detection of D. suzukii Gravid Females and Collection of the First Infested Early Spring Host Fruits Are Concomitant
3.5. Overwintered D. suzukii Females Are Likely the Main Contributors of the Infestation in the First Commercial Fruits
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Asplen, M.K.; Anfora, G.; Biondi, A.; Choi, D.S.; Chu, D.; Daane, K.M.; Gibert, P.; Gutierrez, A.P.; Hoelmer, K.A.; Hutchinson, W.D.; et al. Invasion biology of spotted wing Drosophila (Drosophila suzukii): A global perspective and future priorities. J. Pest Sci. 2015, 88, 469–494. [Google Scholar] [CrossRef]
- De Ros, G.; Conci, S.; Pantezzi, T.; Savini, G. The economic impact of invasive pest Drosophila suzukii on berry production in the Province of Trento, Italy. J. Berry Res. 2015, 5, 89–96. [Google Scholar] [CrossRef]
- Farnsworth, D.; Hamby, K.A.; Bolda, M.; Goodhue, R.E.; Williams, J.C.; Zalom, F.G. Economic analysis of revenue losses and control costs associated with the spotted wing drosophila, Drosophila suzukii (Matsumura), in the California raspberry industry. Pest Manag. Sci. 2017, 73, 1083–1090. [Google Scholar] [CrossRef] [PubMed]
- Goodhue, R.E.; Bolda, M.; Farnsworth, D.; Williams, J.C.; Zalom, F.G. Spotted wing drosophila infestation of California strawberries and raspberries: Economic analysis of potential revenue losses and control costs. Pest Manag. Sci. 2011, 67, 1396–1402. [Google Scholar] [CrossRef] [PubMed]
- De Ros, G.; Anfora, G.; Grassi, A.; Ioriatti, C. The potential economic impact of Drosophila suzukii on small fruits production in Trentino (Italy). IOBC-WPRS Bull. 2013, 91, 317–321. [Google Scholar]
- Cini, A.; Ioriatti, C.; Anfora, G. A review of the invasion of Drosophila suzukii in Europe and a draft research agenda for integrated pest management. Bull. Insectol. 2012, 65, 149–160. [Google Scholar]
- Hauser, M. A historic account of the invasion of Drosophila suzukii (Matsumura) (Diptera: Drosophilidae) in the continental United States, with remarks on their identification. Pest Manag. Sci. 2011, 67, 1352–1357. [Google Scholar] [CrossRef] [PubMed]
- NPPO. First Findings of Drosophila suzukii; Pest Rep.; NPPO: Utrecht, The Netherlands, 2012; p. 2. [Google Scholar]
- Helsen, H.; van der Sluis, B. Drosophila suzukii: Situation in The Netherlands. Unpublished.
- Bruck, D.J.; Bolda, M.; Tanigoshi, L.; Klick, J.; Kleiber, J.; DeFrancesco, J.; Gerdemanc, B.; Spitler, H. Laboratory and field comparisons of insecticides to reduce infestation of Drosophila suzukii in berry crops. Pest Manag. Sci. 2011, 67, 1375–1385. [Google Scholar] [CrossRef] [PubMed]
- Van Timmeren, S.; Isaacs, R. Control of spotted wing drosophila, Drosophila suzukii, by specific insecticides and by conventional and organic crop protection programs. Crop. Prot. 2013, 54, 126–133. [Google Scholar] [CrossRef]
- Rogers, M.A.; Burkness, E.C.; Hutchison, W. Evaluation of high tunnels for management of Drosophila suzukii in fall-bearing red raspberries: Potential for reducing insecticide use. J. Pest Sci. 2016, 89, 815–821. [Google Scholar] [CrossRef]
- Grassi, A.; Gottardello, A.; Dalton, D.T.; Tait, G.; Rendon, D.; Ioriatti, C.; Gibeaut, D.; Rossi Stacconi, M.V.; Walton, V.M. Seasonal Reproductive Biology of Drosophila suzukii (Diptera: Drosophilidae) in Temperate Climates. Environ. Entomol. 2018, 47, 166–174. [Google Scholar] [CrossRef] [PubMed]
- Ioriatti, C.; Boselli, M.; Caruso, S.; Galassi, T.; Gottardello, A.; Grassi, A.; Tonina, L.; Vaccari, G.; Mori, N. Approccio integrato per la difesa dalla Drosophila suzukii. Rivista di Frutticoltura e di Ortofloricoltura 2015, 77, 32–37. [Google Scholar]
- Kenis, M.; Tonina, L.; Eschen, R.; van der Sluis, B.; Sancassani, M.; Mori, N.; Haye, T.; Helsen, H. Non-crop plants used as hosts by Drosophila suzukii in Europe. J. Pest Sci. 2016, 89, 735–748. [Google Scholar] [CrossRef] [PubMed]
- Jones, V.P.; Wiman, N.G. Modeling the interaction of physiological time, seasonal weather patterns, and delayed mating on population dynamics of codling moth, Cydia pomonella (L.) (Lepidoptera: Tortricidae). Popul. Ecol. 2012, 54, 421–429. [Google Scholar] [CrossRef]
- Cianci, D.; van den Broek, J.; Caputo, B.; Marini, F.; Della Torre, A.; Heesterbeek, H.; Hartemink, N. Estimating mosquito population size from mark–release–recapture data. J. Med. Entomol. 2013, 50, 533–542. [Google Scholar] [CrossRef] [PubMed]
- Briem, F.; Eben, A.; Gross, J.; Vogt, H. An invader supported by a parasite: Mistletoe berries as a host for food and reproduction of Spotted Wing Drosophila in early spring. J. Pest Sci. 2016, 89, 749–759. [Google Scholar] [CrossRef]
- Rossi-Stacconi, M.V.; Kaur, R.; Mazzoni, V.; Ometto, L.; Grassi, A.; Gottardello, A.; Rota-Stabelli, O.; Anfora, G. Multiple lines of evidence for reproductive winter diapause in the invasive pest Drosophila suzukii. J. Pest Sci. 2016, 89, 689–700. [Google Scholar] [CrossRef]
- Wang, X.-G.; Stewart, T.J.; Biondi, A.; Chavez, B.A.; Ingels, C.; Caprile, J.; Grant, J.A.; Walton, V.M.; Daane, K.M. Population dynamics and ecology of Drosophila suzukii in Central California. J. Pest Sci. 2016, 89, 701–712. [Google Scholar] [CrossRef]
- Shearer, P.W.; West, J.D.; Walton, V.M.; Brown, P.H.; Svetec, N.; Chiu, J.C. Seasonal cues induce phenotypic plasticity of Drosophila suzukii to enhance winter survival. BMC Ecol. 2016, 16, 11. [Google Scholar] [CrossRef] [PubMed]
- Mitsui, H.; Beppu, K.; Kimura, M.T. Seasonal life cycles and resource uses of flower- and fruit-feeding drosophilid flies (Diptera: Drosophilidae) in central Japan. Entomol. Sci. 2010, 13, 60–67. [Google Scholar] [CrossRef] [Green Version]
- Zerulla, F.N.; Schmidt, S.; Streitberger, M.; Zebitz, C.P.; Zelger, R. On the overwintering ability of Drosophila suzukii in South Tyrol. J. Berry Res. 2015, 5, 41–48. [Google Scholar] [CrossRef]
- Wallingford, A.K.; Lee, J.C.; Loeb, G.M. The influence of temperature and photoperiod on the reproductive diapause and cold tolerance of spotted-wing drosophila, Drosophila Suzukii. Entomol. Exp. Appl. 2016, 159, 327–337. [Google Scholar] [CrossRef]
- Fraimout, A.; Jacquemart, P.; Villarroel, B.; Aponte, D.J.; Decamps, T.; Herrel, A.; Cornette, R.; Debat, V. Phenotypic plasticity of Drosophila suzukii wing to developmental temperature: Implications for flight. J. Exp. Boil. 2018, 221, jeb166868. [Google Scholar] [CrossRef] [PubMed]
- Stockton, D.; Wallingford, A.; Loeb, G. Phenotypic plasticity promotes overwintering survival in a globally invasive crop pest, Drosophila suzukii. Insects 2018, 9, 105. [Google Scholar] [CrossRef] [PubMed]
- Poyet, M.; Le Roux, V.; Gibert, P.; Meirland, A.; Prévost, G.; Eslin, P.; Chabrerie, O. The wide potential trophic niche of the asiatic fruit fly Drosophila suzukii: The key of its invasion success in temperate Europe? PLoS ONE 2015, 10, e0142785. [Google Scholar] [CrossRef] [PubMed]
- Arnó, J.; Solà, M.; Riudavets, J.; Gabarra, R. Population dynamics, non-crop hosts, and fruit susceptibility of Drosophila suzukii in Northeast Spain. J. Pest Sci. 2016, 89, 713–723. [Google Scholar] [CrossRef]
- Lee, J.C.; Dreves, A.J.; Cave, A.M.; Kawai, S.; Isaacs, R.; Miller, J.C.; Van Timmeren, S.; Bruck, D.J. Infestation of wild and ornamental noncrop fruits by Drosophila suzukii (Diptera: Drosophilidae). Ann. Entomol. Soc. Am. 2015, 108, 117–129. [Google Scholar] [CrossRef]
- Tochen, S.; Dalton, D.T.; Wiman, N.; Hamm, C.; Shearer, P.W.; Walton, V.M. Temperature-related development and population parameters for Drosophila suzukii (Diptera: Drosophilidae) on cherry and blueberry. Environ. Entomol. 2014, 43, 501–510. [Google Scholar] [CrossRef] [PubMed]
- Helsen, H.; van der Sluis, B. Egg-to-adult survival of D. suzukii in Aucuba japonica fruits. Unpublished.
- Vonlanthen, O.; Kehrli, P. Caractérisation et apparition des formes hivernale et estivale de Drosophila Suzukii. Revue Suisse de Viticulture, Arboriculture et Horticulture 2015, 47, 318–323. [Google Scholar]
- Robertson, F.W.; Reeve, E. Studies in quantitative inheritance. J. Genet. 1952, 50, 414. [Google Scholar] [CrossRef]
- Cummings, M.; King, R. The cytology of the vitellogenic stages of oogenesis in Drosophila melanogaster. General staging characteristics. J. Morphol. 1969, 128, 427–441. [Google Scholar] [CrossRef]
- Baskerville, G.; Emin, P. Rapid estimation of heat accumulation from maximum and minimum temperatures. Ecology 1969, 50, 514–517. [Google Scholar] [CrossRef]
- Dalton, D.T.; Walton, V.M.; Shearer, P.W.; Walsh, D.B.; Capriled, J.; Isaacs, R. Laboratory survival of Drosophila suzukii under simulated winter conditions of the Pacific Northwest and seasonal field trapping in five primary regions of small and stone fruit production in the United States. Pest Manag. Sci. 2011, 67, 1368–1374. [Google Scholar] [CrossRef] [PubMed]
- Stephens, A.; Asplen, M.; Hutchison, W.; Venette, R.C. Cold hardiness of winter-acclimated Drosophila suzukii (Diptera: Drosophilidae) adults. Environ. Entomol. 2015, 44, 1619–1626. [Google Scholar] [CrossRef] [PubMed]
- Wiman, N.G.; Dalton, D.T.; Anfora, G.; Biondi, A.; Chiu, J.C.; Daane, K.M.; Gerdeman, B.; Gottardello, A.; Hamby, K.A.; Isaacs, R.; et al. Drosophila suzukii population response to environment and management strategies. J. Pest Sci. 2016, 89, 653–665. [Google Scholar] [CrossRef] [PubMed]
- Tochen, S.; Woltz, J.; Dalton, D.; Lee, J.; Wiman, N.; Walton, V. Humidity affects populations of Drosophila suzukii (Diptera: Drosophilidae) in blueberry. J. Appl. Entomol. 2015. [Google Scholar] [CrossRef]
- Tochen, S.; Walton, V.M.; Lee, J.C. Impact of floral feeding on adult Drosophila suzukii survival and nutrient status. J. Pest Sci. 2016, 89, 793–802. [Google Scholar] [CrossRef]
- Kenis, M.; (CABI, Delémont, Switzerland). Sensitivity of Host Fruit Species to Desiccation after Picking. Personal communication. 2018. [Google Scholar]
- Helsen, H.; van der Sluis, B. Field monitoring data on Viscum album in The Netherlands. Unpublished.
- Lee, J.C.; Bruck, D.J.; Curry, H.; Edwards, D.; Haviland, D.R.; Steenwykd, R.A.V.; Yorgey, B.M. The susceptibility of small fruits and cherries to the spotted-wing drosophila, Drosoph Suzukii. Pest Manag. Sci. 2011, 67, 1358–1367. [Google Scholar] [CrossRef] [PubMed]
- Burrack, H.J.; Fernandez, G.E.; Spiveyc, T.; Krausa, D.A. Variation in selection and utilization of host crops in the field and laboratory by Drosophila suzukii Matsumara (Diptera: Drosophilidae), an invasive frugivore. Pest Manag. Sci. 2013, 69, 1173–1180. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.C.; Dalton, D.T.; Swoboda-Bhattarai, K.A.; Bruck, D.J.; Burrack, H.J.; Strik, B.C.; Woltz, J.M.; Walton, V.M. Characterization and manipulation of fruit susceptibility to Drosophila suzukii. J. Pest Sci. 2016, 89, 771–780. [Google Scholar] [CrossRef]
- Wong, J.; Wallingford, A.; Loeb, G.; Lee, J. Physiological status of Drosophila suzukii (Diptera: Drosophilidae) affects their response to attractive odours. J. Appl. Entomol. 2018, 142, 473–482. [Google Scholar] [CrossRef]
- Klick, J.; Yang, W.Q.; Walton, V.M.; Dalton, D.T.; Hagler, J.R.; Dreves, A.J.; Lee, J.C.; Bruck, D.J. Distribution and activity of Drosophila suzukii in cultivated raspberry and surrounding vegetation. J. Appl. Entomol. 2016, 140, 37–46. [Google Scholar] [CrossRef]
- Klick, J.; Yang, W.Q.; Bruck, D.J. Marking Drosophila suzukii (Diptera: Drosophilidae) with Rubidium or 15N. J. Econ. Entomol. 2015, 108, 1447–1451. [Google Scholar] [CrossRef] [PubMed]
- Klick, J.; Lee, J.C.; Hagler, J.R.; Bruck, D.J.; Yang, W.Q. Evaluating Drosophila suzukii immunomarking for mark-capture research. Entomol. Exp. Appl. 2014, 152, 31–41. [Google Scholar] [CrossRef]
- Tait, G.; Grassi, A.; Pfab, F.; Crava, C.M.; Dalton, D.T.; Magarey, R.; Ometto, L.; Vezzulli, S.; Rossi-Stacconi, M.V.; Gottardello, A. Large-scale spatial dynamics of Drosophila suzukii in Trentino, Italy. J. Pest Sci. 2018. [Google Scholar] [CrossRef]
- Ryan, G.D.; Emiljanowicz, L.; Wilkinson, F.; Kornya, M.; Newman, J.A. Thermal Tolerances of the Spotted-Wing Drosophila Drosophila suzukii (Diptera: Drosophilidae). J. Econ. Entomol. 2016, 109, 746–752. [Google Scholar] [CrossRef] [PubMed]
- Thistlewood, H.M.; Gill, P.; Beers, E.H.; Shearer, P.W.; Walsh, D.B.; Rozema, B.M.; Acheampong, S.; Castagnoli, S.; Yee, W.L.; Smytheman, P. Spatial analysis of seasonal dynamics and overwintering of Drosophila suzukii (Diptera: Drosophilidae) in the Okanagan-Columbia basin, 2010–2014. Environ. Entomol. 2018, 47, 221–232. [Google Scholar] [CrossRef] [PubMed]
- Partridge, L.; Barrie, B.; Barton, N.H.; Fowler, K.; French, V. Rapid laboratory evolution of adult life-history traits in Drosophila melanogaster in response to temperature. Evolution 1995, 49, 538–544. [Google Scholar] [CrossRef] [PubMed]
- Nikolouli, K.; Colinet, H.; Renault, D.; Enriquez, T.; Mouton, L.; Gibert, P.; Sassu, F.; Cáceres, C.; Stauffer, C.; Pereira, R. Sterile insect technique and Wolbachia symbiosis as potential tools for the control of the invasive species Drosophila suzukii. J. Pest Sci. 2017, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Girod, P.; Borowiec, N.; Buffington, M.; Chen, G.; Fang, Y.; Kimura, M.T.; Peris-Felipo, F.J.; Ris, N.; Wu, H.; Xiao, C. The parasitoid complex of D. suzukii and other fruit feeding Drosophila species in Asia. Sci. Rep. 2018, 8, 11839. [Google Scholar] [CrossRef] [PubMed]
- Girod, P.; Lierhmann, O.; Urvois, T.; Turlings, T.C.; Kenis, M.; Haye, T. Host specificity of Asian parasitoids for potential classical biological control of Drosophila suzukii. J. Pest Sci. 2018, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Daane, K.M.; Wang, X.G.; Biondi, A.; Miller, B.; Miller, J.C.; Riedl, H.; Shearer, P.W.; Guerrieri, E.; Giorgini, M.; Buffington, M.; et al. First exploration of parasitoids of Drosophila suzukii. J. Pest Sci. 2016, 89, 823–835. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Species | Jan | Feb | Mar | Apr | May | Jun | Jul | Aug | Sep | Oct | Nov | Dec |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Aucuba japonica | ||||||||||||
| Elaeagnus x ebbingei | ||||||||||||
| Skimmia japonica | ||||||||||||
| Hedera helix | ||||||||||||
| Viscum album | ||||||||||||
| Cherry crops | ||||||||||||
| Soft fruit crops | ||||||||||||
| Grape crops |
| Plant Species | No. Sampled Locations | No. Collected Fruits | No. D. suzukii Adults Reared | Infestation Rate (%) |
|---|---|---|---|---|
| Cotoneaster spp. | 2 | 470 | 0 | 0.0 |
| Elaeagnus x ebbingei | 1 | 758 | 38 | 5.0 |
| Hedera helix | 4 | 550 | 0 | 0.0 |
| Aucuba japonica | 7 | 791 | 61 | 7.7 |
| Skimmia japonica | 1 | 860 | 6 | 0.7 |
| Viscum album | 1 | 1870 | 55 | 2.9 |
| No. Collected Fruits | % Infested Fruits | Incubation in the Laboratory | |||||
|---|---|---|---|---|---|---|---|
| No. Infested Fruits 1 | No. Eggs | No. Emerged Flies | Hatch Rate | ||||
| 17 April | Aucuba japonica | 282 | 35.1 | 48 | 87 | 3 | 3.4 |
| Elaeagnus x ebbingei | 213 | 2.3 | 5 | 5 | 0 | 0.0 | |
| Skimmia japonica | 308 | 1.0 | 3 | 6 | 0 | 0.0 | |
| Hedera helix | 1472 | 0.2 | 3 | 3 | 0 | 0.0 | |
| Viscum album | 374 | 0.0 | 0 | 0 | 0 | 0.0 | |
| 8 May | Aucuba japonica | 321 | 53.0 | 97 | 189 | 20 | 10.6 |
| Elaeagnus x ebbingei | 322 | 10.9 | 35 | 43 | 16 | 37.2 | |
| Skimmia japonica | 347 | 0.9 | 3 | 3 | 0 | 0.0 | |
| Hedera helix | 1400 | 0.2 | 3 | 3 | 0 | 0.0 | |
| Viscum album | 412 | 0.0 | 0 | 0 | 0 | 0.0 | |
| Field Incubation | Laboratory Incubation | |||||||
|---|---|---|---|---|---|---|---|---|
| Incubation Date 1 | No. Infested Fruits | No. Eggs | No. Emerged Adults | Hatch Rate | No. Infested Fruits | NO. EGGS | No. Emerged Adults | Hatch Rate |
| 10 April | 25 | 40 | 0 | 0.0 | 2 | 40 | 1 | 2.5 |
| 17 April | 49 | 94 | 0 | 0.0 | 48 | 87 | 3 | 3.4 |
| 23 April | 63 | 132 | 2 | 1.5 | 62 | 128 | 4 | 3.1 |
| 1 May | 73 | 158 | 5 | 3.2 | 70 | 149 | 4 | 2.7 |
| 8 May | 73 | 101 | 5 | 5 | 97 | 189 | 20 | 10.6 |
| 15 May | 135 | 340 | 42 | 12.4 | 127 | 323 | 84 | 26 |
| 22 May | 121 | 441 | 10 | 2.3 | 116 | 414 | 15 | 3.6 |
| 29 May | 79 | 73 | 0 | 0 | 179 | 81 | 0 | 0 |
| Total | 618 | 1379 | 64 | 4.6 | 724 | 1411 | 131 | 9.3 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Panel, A.D.C.; Zeeman, L.; Van der Sluis, B.J.; Van Elk, P.; Pannebakker, B.A.; Wertheim, B.; Helsen, H.H.M. Overwintered Drosophila suzukii Are the Main Source for Infestations of the First Fruit Crops of the Season. Insects 2018, 9, 145. https://doi.org/10.3390/insects9040145
Panel ADC, Zeeman L, Van der Sluis BJ, Van Elk P, Pannebakker BA, Wertheim B, Helsen HHM. Overwintered Drosophila suzukii Are the Main Source for Infestations of the First Fruit Crops of the Season. Insects. 2018; 9(4):145. https://doi.org/10.3390/insects9040145
Chicago/Turabian StylePanel, Aurore D. C., Laura Zeeman, Bart J. Van der Sluis, Peter Van Elk, Bart A. Pannebakker, Bregje Wertheim, and Herman H. M. Helsen. 2018. "Overwintered Drosophila suzukii Are the Main Source for Infestations of the First Fruit Crops of the Season" Insects 9, no. 4: 145. https://doi.org/10.3390/insects9040145