
Abstract
Key message
The method of graphical genotyping is applied to a panel of tetraploid potato cultivars to visualize haplotype sharing. The method allowed to map genes involved in virus and nematode resistance. The physical coordinates of the amount of linkage drag surrounding these genes are easily interpretable.
Abstract
Graphical genotyping is a visually attractive and easily interpretable method to represent genetic marker data. In this paper, the method is extended from diploids to a panel of tetraploid potato cultivars. Application of filters to select a subset of SNPs allows one to visualize haplotype sharing between individuals that also share a specific locus. The method is illustrated with cultivars resistant to Potato virus Y (PVY), while simultaneously selecting for the absence of the SNPs in susceptible clones. SNP data will then merge into an image which displays the coordinates of a distal genomic region on the northern arm of chromosome 11 where a specific haplotype is introgressed from the wild potato species S. stoloniferum (CPC 2093) carrying a gene (Ny (o,n)sto ) conferring resistance to two PVY strains, PVYO and PVYNTN. Graphical genotyping was also successful in showing the haplotypes on chromosome 12 carrying Ry-f sto , another resistance gene derived from S. stoloniferum conferring broad-spectrum resistance to PVY, as well as chromosome 5 haplotypes from S. vernei, with the Gpa5 locus involved in resistance against Globodera pallida cyst nematodes. The image also shows shortening of linkage drag by meiotic recombination of the introgression segment in more recent breeding material. Identity-by-descent was found to be a requirement for using graphical genotyping, which is proposed as a non-statistical alternative method for gene discovery, as compared with genome-wide association studies. The potential and limitations of the method are discussed.
Similar content being viewed by others

Introduction
The concept of graphical genotypes as a visual method to represent genetic marker data was first described by Young and Tanksley (1989). In a karyotype-style drawing, they used colours to display the mosaic structure of the chromosomes in F2 or backcross individuals according to the parental origin of the chromosomal regions or alleles. The transition of one colour into another displayed how chromosomes were transmitted and indicated the positions of crossovers that occurred during the meiosis of the F1 plants. Graphical genotypes are visually attractive and more easily interpretable by glance than the numerical information in a spreadsheet with offspring genotypes in columns and marker data in rows.
Graphical genotypes are used for a number of analyses. First, graphical genotypes can display the positions and the proportions of donor and recurrent genome during subsequent backcross generations (Young and Tanksley 1989; Yun et al. 2006). Second, graphical genotypes allow quick data inspection and the visual identification of singletons (Fig. 1a) that should not be interpreted as double recombinants (Van Os et al. 2005). Good data quality should result in a “Zebra striping pattern”, whereas erroneous data add isolated spots. Thirdly, high-resolution mapping of loci with major effects (Finkers-Tomczak et al. 2009) is supported by a graphical analysis of the recombinants and their trait values, allowing one to zoom into the remaining interval where candidate genes reside. Graphical genotyping is supported by software packages such as GGT (van Berloo 1999, 2008) and Flapjack (Milne et al. 2010).

Two hypothetical examples of graphical genotypes. a Graphical genotypes of a bi-parental diploid backcross offspring where colour codes distinguish the parental origin of the chromosomal segment (red indicates the recurrent parent, yellow indicates heterozygous introgression of donor parent). Singletons, putatively indicating error data, are indicated with the black circles. b Graphical genotypes of unrelated tetraploid cultivars, where the dosage of the minor allele is indicated with numbers 0, 1, 2, 3 and 4 or the shades red, yellow, light green, middle green and dark green (color figure online)
Usually, graphical genotypes are generated using offspring from bi-parental crosses where two colours can encode the contribution of each of the two parents. A third colour may be needed to depict heterozygosity, but in BC1 or recombinant inbred lines (RILs) a third colour is not necessary. The F1 offspring from non-inbred parents is cumbersome, because four colours are required to show the mosaic structure in the maternally and paternally inherited recombinant chromosomes. This is easily circumvented by the separation of maternal and paternal markers and to apply graphical genotyping separately to the parental maps. An example is shown in Fig. 1a, which could represent a parental map of non-inbred parents, doubled haploids (DHs), RILs or a BC1.
Graphical genotyping of unrelated clones may be possible for diploid, self-pollinated species. Here, the graphical image can be used to display a local haplotype map (Motte et al. 2014) where DNA sequence variants representing either the allele of the reference genome or the non-reference allele are shown with different colour codes. Graphical genotypes and local haplotype maps are seemingly different terms (used in a mapping or haplotyping context, respectively), but they are conceptually related. Multi-colour graphical genotyping is used for depicting multiparent advanced generation inter-cross (MAGIC) lines (Bergelson and Roux 2010). Graphical genotyping of a panel of unrelated tetraploids seems to be without value, because no meaningful pattern is displayed (Fig. 1b).
Potato virus Y (PVY) is the most widespread and economically harmful virus infecting potatoes. It can cause 50–85% yield reduction (Valkonen 2007). PVY is transmitted by aphids and difficult to control in the field unless resistant potato cultivars are grown. Although many countries have implemented systems for the propagation of virus-free planting material, breeding for PVY resistance offers a more durable solution. Resistance genes against PVY have been introgressed in potato cultivars from many sources, as summarized in Table 1.
Solanum stoloniferum is an important source of PVY resistance (Ross 1986). It carries genes for extreme resistance (ER) able to inhibit multiplication of all strains of PVY. Two genes, Ry-f sto and Ry sto , for ER to PVY derived from S. stoloniferum have been mapped to the same region on chromosome 12 and used in variety breeding. Ry sto tends to be associated with male sterility, in contrast to Ry-f sto (Flis et al. 2005; Song and Schwarzfischer 2008). Brigneti et al. (1997) reported on the localisation of a PVY resistance from S. stoloniferum on chromosome 11, but their work has not received much follow-up and their locus name Ry sto has caused confusion due to synonymy with the gene subsequently described on chromosome 12. Brigneti et al. (1997) used clone I-1039, which was developed in India as cv. KHUMAL RED 2 from Scottish late blight-resistant progenitor clones and an undisclosed progenitor M 136-6. This material has Solanum phureja and S. edinense in its pedigree, but its S. stoloniferum origin is unclear from pedigree information presented by the authors. Clone I-1039 is rarely used in breeding and resulted in two Danish cultivars TIVOLI and LIVA, and two cultivars released in Ecuador and Rwanda, FRIPAPA 99 and GIKUNGU, respectively. Hence, it may be problematic to extrapolate the marker data of Brigneti et al. (1997), for the identification of PVY resistance beyond their experimental mapping population. Another donor of PVY resistance, clone F87084, derived from S. stoloniferum CPC 2093 (De Jong et al. 2001; Nie et al. 2015) has not yet resulted in cultivars released to the market and its map position is not described. Resistance to PVY originating in S. stoloniferum has been introgressed also via three German progenitor clones: MPI 13128 (sto × ERICA), clone 43 (sto × POLONIA), MPI 46.152/1 (sto × FRÜHMOLLE) (Song and Schwarzfischer 2008), and from the Commonwealth Potato Collection (accession CPC 2093), e.g. via the Dutch progenitor clone Y 66-13-636.
In this study, we applied graphical genotyping on unrelated tetraploid potato cultivars as a tool for mapping PVY resistance and identification of the introgressed haplotypes. The aim was to elucidate whether the Dutch potato cultivars descending from S. stoloniferum CPC 2093 via progenitor clone Y 66-13-636 contain a locus for resistance to PVY on chromosome 11. Furthermore, we tested whether this method allows identification of introgression segments of the Gpa5 locus (Rouppe van der Voort et al. 2000) and the Ry-f sto locus (Song et al. 2005; Flis et al. 2005). Finally, we also investigated whether other traits lacking most recent common ancestor (MRCA) information can be identified using this method.
Materials and methods
Plant material and PVY resistance tests
A panel of 83 tetraploid cultivars and progenitor clones were included in the study (Supplementary data S1). Among these tetraploids, the authors were aware that cultivar FESTIEN is resistant to PVY, possibly due to a resistance gene derived from its great-grandparent Y 66-13-636. KARTEL, the PVY-susceptible parent of FESTIEN, was also included in the panel. Pedigree information was retrieved from our database (Berloo et al. 2007).
Validation of the PVY resistance gene was achieved by growing 180 offspring of the cross between FESTIEN (PVY resistant) × SERESTA (PVY susceptible) in pots in the greenhouse and inoculating them by hand with a PVY tuber necrosis strain (PVYNTN) 4 weeks after planting. Two weeks later, symptoms of primary infection in inoculated leaves were scored. All 180 genotypes were analysed by ELISA using monoclonal antiserum (obtained from Plant Research International, Wageningen, The Netherlands). The plants negative for PVY in ELISA and without symptoms were defined as resistant, whereas the remaining plants were defined as susceptible.
FESTIEN was characterized for the type of PVY resistance using two well-defined PVY strains representing the PVY strain group O (isolate PVYO-UK; GenBank accession no. JX424837) (Tian and Valkonen 2013) and PVY strain group N (isolate PVYN-NTN-Nevski; JX432967) (Tian et al. 2014). Potato virus A (isolate PVA-U; AJ131402) (Rajamäki et al. 1998) was also included in the experiments. The viruses were maintained in potato cv. PITO. Healthy plants of FESTIEN were multiplied by taking and rooting stem cuttings in a growth chamber under 16-h photoperiod, light intensity 250 μE m−2 s−1, temperature 23/20 °C day/night. The plants were watered as needed and fertilized with 1% fertilizer (N:P:K = 16:9:22, Yara, Espoo, Finland). Plants were side-graft-inoculated with PVY-O, PVY-N or PVA, three plants per virus, and monitored weekly for symptom development in two independent experiments. In both experiments, the uppermost fully expanded leaves were sampled for the first time at 25 and 30 days post-inoculation (dpi), respectively, monitored further for 30 and 40 days, respectively, and sampled again. The sampled leaves were tested by DAS-ELISA using polyclonal antibodies to PVY (Adgen, Auchincruive, Ayr, Scotland, UK) and monoclonal antibodies to PVA (Mab 58/0) (Rajamäki et al. 1998). Leaf samples were weighed, 1 g of tissue was ground in 3 ml of ELISA sample buffer, and two aliquots (100 µl) of the sap were transferred to two wells of the microtitre plate (Greiner Laborteknik, Frickenhausen, Germany). Leaf sap from the potato plants used as sources of virus for inoculation was included as positive controls. Colour reactions were developed using p-nitrophenyl phosphate as a substrate, and absorbance was recorded at 405 nm using a microtitre plate reader (Benchmark, Bio-Rad, Hercules, CA, USA). ELISA absorbance values (A 405) were regarded as positive if they were two times higher (±standard deviation) than those of the healthy control plants (Fenlon and Sopp 1991).
DNA sequence variants
The sequence of the reference genome of Solanum tuberosum Group Phureja DM1-3 516R44 (DM) was obtained from the Potato Genome Sequencing Consortium (PGSC 2011). The order of superscaffolds (referred to as DMB followed by the scaffold number) is according to version 4.03 and can be retrieved from the study by Sharma et al. (2013). Next-generation sequencing of a panel of 83 tetraploid cultivars allowed the identification of 129,156 DNA sequence variants (Uitdewilligen et al. 2013). Here, the relative read depth at variant positions was used for tetraploid genotype calling in the cultivars. Further details about the methods that resulted in this dataset can be retrieved from the paper by Uitdewilligen et al. (2013). A DNA sequence variant from this study may refer to a SNP, a multinucleotide polymorphism (MNP), indels, and/or multi-allelic SNPs, hereafter for brevity collectively referred to as SNPs.
Genotype calling can be performed using the following terms: the reference (DM) and alternative (non-DM) allele, or the minor and major allele, abbreviated hereafter as REF and ALT or MIN and MAJ. Here, genotype calls representing the ALT dosage values indicate that a genotype has 0, 1, 2, 3 or 4 copies of the non-DM allele as compared with the DM reference (REF) genome. The cultivar panel of 83 tetraploids represents a population of 332 alleles (4 × 83) and for each DNA variant its allele frequency has been calculated as the sum of the copies per cultivar divided by 332. Hence, population allele counts ranging from 1 to 166 result in a minor allele frequency (MAF), and the allele with such an allele frequency equal or below 50% is considered as the minor allele (MIN). Allele counts ranging from 167 to 331 result in an allele frequency >50% and such alleles are regarded as the major allele (MAJ).
The colour codes are not defined relative to the DM reference genome. If the DM reference genome has a sequence variant which is quite rare in the remainder of the gene pool then the DM reference genome represents the minor allele.
Filtering of SNPs dosage data to construct graphical genotypes
The data were loaded in Microsoft Excel where the 83 cultivars are shown in columns and the sequence variants along with their coordinates in 129,156 rows. The rows were sorted according to chromosome, superscaffold order and coordinates on pseudomolecules version 4.03 (Sharma et al. 2013). In the header row of the spreadsheet, we inserted filters and used these as logical operators (e.g. ‘IF’, ‘AND’, or ‘NOT’ statements). Specific allele frequencies can be filtered by setting the ‘number filter’ and choosing ‘is smaller than’. To remove rows with SNPs ‘NOT’ involved in, e.g. resistance, a ‘number filter’ was set at ‘is equal to zero’. The combination of setting additional filters on multiple columns (varieties) will act as the ‘AND’ operator. Unfortunately, the ‘OR’ operator to combine criteria across columns is not offered in Excel.
To select haplotypes or chromosomal regions involved in PVY resistance, the filters were set to select SNPs with a small allele frequency ‘AND’ specific minor alleles should be present in FESTIEN and Y 66-13-636 (allele dosage ≥1). For columns representing susceptible cultivars, the filter was set at zero for the minor allele, which selects for nulliplex genotypes. Graphical genotypes were displayed in colour using conditional formats in Excel where pseudo-colours indicated allele dosage per genotype ranging from red to green for nulliplex to quadruplex genotypes, respectively.
GWAS
The above analysis was verified with a statistical analysis. For this purpose, trait values were defined for 83 potato cultivars (trait values are resistant = 1; susceptible = 0) and a naive genome-wide association study (GWAS) was performed using GenStat 15th edition (VSN International Ltd, UK).
Results
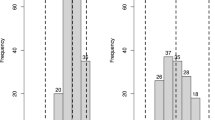
When SNP data were loaded in Excel, conditional formatting according to colour scales was set as shown in Fig. 1. This resulted in a predominant background colour with rows containing fewer or greater numbers of cells having the alternative pseudo-colour according to the population allele frequency. This allows the interpretation that the vast majority of the SNPs alleles represent rare haplotypes. Figure 2 displays a clear L-shaped distribution of the allele frequencies of the SNPs. This implies that if SNPs were phased into haplotypes, the haplotype allele frequency will also display such an L-shaped distribution, which suggests that the potato gene pool is composed of many low-frequency haplotypes.

The distribution of the minor allele frequencies of all 129,156 sequence variants. The distribution indicates that many SNPs are rare variants (after Uitdewilligen et al. 2013)
Filtering of SNPs data was performed in Excel with three criteria. As PVY resistance must have a low allele frequency, the first filter for the column with the allele count was set below 20 (allele frequency of ~6.0%). This reduced the list of 129K SNPs to 58K. The second and third filter were set to ≥1, to select for SNPs having at least ≥1 minor alleles in the column of the two PVY-resistant clones FESTIEN ‘AND’ Y 66-13-636. These clones must carry at least one allele conferring PVY resistance. The combined criteria for allele frequency ‘AND’ present in two varieties greatly reduced the number of SNPs from 58K to 748 SNPs. The next filters in the columns of PVY-susceptible clones, such as KARTEL (the susceptible parent of FESTIEN), KATAHDIN, BINTJE and ARRAN PILOT (old cultivars), were set to 0, indicating the absence of an allele involved in resistance. This selection against SNPs alleles from these four susceptible varieties resulted in only 531 SNPs matching all aforementioned criteria, which represent only 0.41% of all SNPs. Interestingly, with conditional formats these SNP data produced a specific striping pattern as shown in Fig. 3. This pattern is indeed reminiscent of graphical genotypes as observed for diploid bi-parental F2 or backcross populations. However, the cultivar panel is comprised of highly diverse material. If any haplotype was shared between members of the cultivar panel, then haplotype-specific SNP alleles (hs-SNPs) would merge into a yellow vertical bar in the graphical genotyping image.

Graphical genotyping of 83 tetraploid cultivars (columns) with 531 SNPs (in rows) from 12 potato chromosomes. Absence or presence of minor alleles is indicated by conditionally formatted cells in red (0) or yellow to green (1, 2, 3, 4), respectively. Settings for allele dosages in FESTIEN and Y 66-13-636 were ≥1 (yellow to green columns, depending on allele dosage), whereas KARTEL, KATAHDIN, BINTJE and ARRAN PILOT were set <1 (red columns). The apparent pattern is indicative of haploblock sharing. The haploblock of chromosome 11 indicates the introgression segment that carries the PVY resistance gene descending from S. stoloniferum CPC2093 (color figure online)
Haplotype blocks could be identified for regions that belong to potato chromosome 4, 5, 7, 10 and 11. For all regions, except the region on chromosome 11, the yellow patterns are also observed in potato cultivars that are susceptible. Based on the many false positives these regions were rejected. The candidate region on chromosome 11 indicated that one haploblock was shared by three cultivars: EOS, FESTIEN and Y 66-13-636. This prompted us to examine the phenotype and pedigree of EOS. Indeed, EOS appeared to be highly resistant against PVY, male fertile (personal communication with Jacob Eising, potato breeder at Den Hartigh B.V.), and has Y 66-13-636 as grandfather. Therefore, EOS is not a false positive but a third member of the cultivar panel with PVY resistance derived from S. stoloniferum.
Size of the haploblocks
The map location and sizes of the haploblocks on potato chromosome 4, 5, 6, 7, 10 and 11 (Fig. 3) can be deduced from the physical coordinates of the first and last SNP in the block. For chromosome 4, the haploblock is part of the peri-centromeric heterochromatin and is roughly 10 Mb long, ranging from DMB366 to DMB13 (PGSC 4.03 coordinates chr04:21464000..31110000). For chromosome 5, the haploblock is part of the north arm and is only 1.1 Mb long, ranging from DMB51 to DMB424 (PGSC 4.03 coordinates chr05:4250427..5369289). For chromosome 6, the haploblock with 12 SNPs, locates on the distal end of the south arm in scaffold DMB686 chr06:57791621..57992915. For chromosome 7, the SNPs were found on two scaffolds, both on the north (short) arm. One block is part of the 2.2 Mb most distal scaffold DMB47 and one block is 8 Mb further on the 0.2 Mb scaffold DMB684. For chromosome 10, the 1 Mb haploblock on the north arm is part of two adjacent scaffolds DMB599 and DMB338 (PGSC 4.03 coordinates chr10:3903000..4917000). For chromosome 11, all SNPs of the haploblock belong to the most distal scaffold DMB148 on the north arm (PGSC 4.03 coordinates chr11:1..1439384). The first and the last SNP of the haploblock are 500 kb apart having coordinates chr11:284162 and chr:814554, respectively. This is approximately 500 kb from the well-known R gene cluster that includes genes homologous to N and N-like (Nl-25) TMV resistance genes and analogues (Hehl et al. 1999).
The haploblock comprising PVY resistance from S. stoloniferum CPC 2093
Further refinement of the chromosome 11 haploblock (Fig. 3) comprising PVY resistance from S. stoloniferum CPC 2093 was performed to identify accurate hs-SNPs using more strict filter settings, where the allele count was set to ≤4 (allowing at most one genotyping error), and EOS, FESTIEN and Y 66-13-636 have an allele count ≥1. This resulted in a total of 65 SNPs (Supplementary data 2) which displayed the minor allele in simplex condition in EOS, FESTIEN and Y 66-13-636 and was nulliplex in the other members of the cultivar panel. These SNPs should allow marker-assisted selection in any genetic background without the risk of false-positive results. From these 65 SNPs, a subset of seven SNPs was selected for validation. Their identity and coordinates are as follows: PotVar0063974 (Chr11:284168) a [T/C] SNP, PotVar0064044 (Chr11:398248) a [T/C] SNP, PotVar0064080 (Chr11:398597..398598) a dinucleotide polymorphism [TT/GA], PotVar0064400 (Chr11:786626) a [G/A] SNP, PotVar0064470 (Chr11:787325) a [C/A] SNP, PotVar0064502 (Chr11:787571) a [C/T] SNP, and PotVar0064578 (Chr11:809990) a [T/C] SNP, all from DMB148. The mapping of PVY resistance to DMB148 suggests that the R gene is a member of a well-known R gene cluster, which was named cluster XIa-TNL in bin4-8 (Bakker et al. 2011), or cluster 49 (Jupe et al. 2012). The seven SNPs were selected to be free (as much as possible) from flanking SNPs to avoid assay failure, and were included in a custom-made 20K Infinium SNP array (Vos et al. 2015), used to analyse a much wider panel of 537 potato cultivars described by D’hoop et al. (2014). In this panel, the seven SNPs were validated, because all SNPs identified other descendants from Y 66-13-636: CUPIDO, CYCLOON, LADY CHRISTL, LADY FELICIA, MELODY, MUSICA, ORCHESTRA, ORIANA, SAVIOLA, SANTÉ and W 72-22-496, as well as MIRAKEL, descending from Y 62-2-221 (the resistant parent of Y 66-13-636). These cultivars are simplex for the minor allele and their PVY resistance is in agreement with phenotypic data from breeders’ websites. In susceptible clones, however, the SNPs identified only the major reference allele.
The Infinium array also enabled the identification of four cultivars: ARIZONA, BELANA, OSIRA, and SAGITTA which are positive for this introgression segment, but bear an unknown genetic relationship to CPC 2093. Hence, we predict that these cultivars are resistant, which was verified at the breeders’ website. Furthermore, ALTUS, AXION, CYRANO, DONALD, SERESTA and XANTIA were negative for the SNPs, although these cultivars descend from CPC2093 (Supplementary data 3). It is concluded that these clones no longer carry the introgression segment and are most likely PVY susceptible.
Validation of the map position and phenotypic characterisation of Ny(o,n)sto
The locus involved in PVY resistance as identified using a panel of distantly related cultivars was validated with a new bi-parental mapping population descending from FESTIEN × SERESTA. In this mapping population, 81 individuals were found to be susceptible, whereas 98 were resistant (no symptoms, and no virus detected by ELISA, Supplementary data 4). This is in agreement with a 1:1 segregation ratio and in accordance with the Mendelian expectations of a simplex allele. The two [T/C] SNP markers PotVar63973 (Chr11:284162) and PotVar63974 (Chr11: 284168) were converted into one SNP assay and analysed for co-segregation with the PVY resistance. The SNP assay nicely predicted the PVY resistance phenotype in the offspring of 179 individuals, except for four resistant individuals. This may indicate recombination between the R gene and the marker locus PotVar63973/63974 in four progeny plants, but the lack of infection might also be due to unsuccessful inoculation. The latter is more likely because in the wider panel of 537 cultivars tested with the Infinium array no recombination events were observed among the SNP loci. These results provide further evidence for the localisation of the PVY resistance gene at a distal position of potato chromosome 11.
Further characterisation of PVY resistance in FESTIEN was carried out by graft inoculation with PVYO-UK and PVYN-NTN-Nevski. Both PVY strains induced similar symptoms of necrotic pinpoint lesions in the uppermost leaves 3 weeks post graft inoculation. Necrosis advanced by time and affected also the small veins. Hence, the top leaves became heavily diseased by 6 weeks post inoculation, but they did not die. The plant continued to develop new leaves and these leaves developed similar symptoms (Fig. 4). The plants were stunted, as the top leaves did not develop normally. FESTIEN was also tested for resistance to PVA by graft inoculation, which caused severe systemic necrosis, typical of the hypersensitive resistance response (HR). Initially, veinal necrosis was observed in the top leaves ca. 3 weeks post graft inoculation, followed by development of large necrotic lesions and, eventually, death of the leaf (Fig. 4). Both PVY strains and PVA could be detected in the symptomatic leaves by DAS-ELISA, but virus concentrations in FESTIEN were at least 25–100-fold lower (1–4%) than in the susceptible cultivar PITO. Because the viruses were readily detectable by ELISA in the symptomatic leaves, albeit at very low concentration, FESTIEN was concluded to express HR to both strains of PVY and to PVA. Based on the HR phenotype, we propose Ny (o,n)sto as name for this locus.

Symptoms caused by systemic infection with Potato virus Y (PVY) or Potato virus A (PVA) in the top leaves of potato cv. FESTIEN 3 weeks after side-graft inoculation of the plants. a A healthy leaf from a non-inoculated control; b, c, Necrotic pinpoint lesions caused by the strain PVYNTN-Nevski and PVYO-UK, respectively. d Rapidly expanding necrosis in the leaves systemically infected with PVA
Results of GWAS
A genome-wide association study was performed as a statistical alternative to the graphical genotyping method to analyse the significance of the mapping and the power of GWAS to identify the PVY resistance based on only four resistant cultivars (EOS, FESTIEN, KURAS and Y 66-13-636) in the panel of 83 tetraploids. We included cv. KURAS as the fourth PVY-resistant clone to mimic the situation that GWAS is based on phenotypic data and does not have a priori assumptions on trait heterogeneity (=Variation caused by different genes can give rise to the same phenotype). Figure 5 shows a Manhattan plot with the outcome of the GWAS for Y-virus resistance. A major peak can be observed for SNPs that reside in superscaffold DMB148 on chromosome 11, matching with the coordinates of hs-SNPs in the graphical genotypes. Figure 5 illustrates that a haplotype tagged by several hs-SNPs creates a pileup in the Manhattan plot, whereas spurious or singleton SNPs were detected on chromosomes 2, 3, 6, 7 and 12. The SNPs underlying the peak on the right end of chromosome 12 reside in superscaffold DMB114 and represent the haplotype that carries Ry-f sto (Song and Schwarzfischer 2008). The peak on the right end of chromosome 6 represents a false-positive QTL, based on the coincidental presence of two partial and two complete haplotype blocks in four varieties ARROW, EOS, FESTIEN and Y66-13-636 (see the graphical genotyping image Fig. 3), where ARROW is not descending from CPC 2093. In a GWAS, where KURAS is excluded from the analysis, the secondary peak on chromosome 12 is absent (data not shown).

Manhattan plot of a GWAS, showing the statistical validation of the graphical mapping of Ny (o,n)sto on superscaffold DMB148 on potato chromosome 11. A secondary peak on chromosome 12 is indicative of Ry-f sto . On the y-axis the −log10(P) value, and on the x-axis the physical positions of the SNP is shown in alternating colours for 12 consecutive potato chromosomes
Graphical genotyping of Ry-fsto
We tested if graphical genotyping could also identify the Ry-f sto locus. In our cultivar panel, the variety KURAS was the only genotype identified as being PVY resistant due to Ry-f sto . A value for allele count was set <2, and a value for a minor allele ≥1 for KURAS. This results in the identification of each of the 479 KURAS-specific SNPs as potentially associated with the Ry-f sto haplotype. However, these SNPs were located on segments spanning the entire genome. Scaffold DMB127 on chromosome 3 contributed 75 SNPs (16%); twelve scaffolds of chromosome 7 contributed 321 SNPs (67%), and 50 SNPs (10%) localized to chromosome 12. Within chromosome 12 the SNPs localized at three positions: 11 SNPs telomeric north arm, 13 SNPs close to the centromere and telomeric south we observed 2 SNPs in DMB38 and adjacently 24 SNPs in DMB114 (length 1.6 Mb, coordinates chr12:58927871..60583523). This is indeed the R gene cluster tagged by STM0003 (coordinates: chr12:60055231..60055365; Song et al. 2005) and the PCR markers YES3-3A (genbank accession BV725480; BLAST hit on coordinates: chr12:59061649..59061903).
Nevertheless, this result is incorrect. Three of the SNPs from this introgression segment were present on our 20K-SNP array, but were negative for AMADO, a PVY-resistant descendant of KURAS that was present in the wider panel of 537 potato cultivars. However, other SNPs on the Infinium array could accurately predict PVY resistance based on Ry-f sto . Therefore, graphical genotyping was performed with more relaxed settings. When the filter for allele count was set ≤5 an unexpected haplotype sharing between cv. KURAS and cv. HINDENBURG could be observed in the graphical genotyping image. From other data, we already knew that our DNA was isolated from a clone incorrectly labelled as HINDENBURG.
A second attempt included this incorrectly labelled clone (presumably carrying Ry-f sto ) along with the additional knowledge on the location of Ry-f sto on DMB114, chromosome 12. This resulted in the identification of 62 SNPs (Supplementary data 5). Four of these 62 SNPs (PotVar0052353 at DMB114:626106, PotVar0052707 at DMB114:902759, PotVar0053235 at DMB114:1376622 and PotVar0053451 at DMB114:1599751) were present on the 20K Infinium array and all four SNPs positively identified KURAS and AMADO and were negative for the remainder of the 537 cultivar panel.
The Solanum vernei introgression segment with Gpa5
Another example of the application of graphical genotyping relates to the Gpa5 vrn locus on chromosome 5 conferring Globodera pallida nematode resistance, initially described using the diploid clone 3704-76 (Rouppe van der Voort et al. 2000). The resistant parental clones 3778-16 and 3704-76 are dihaploids extracted from the tetraploid progenitor clones AM 78-3778 and AM 78-3704, respectively. Both clones may carry multiple S.vernei-derived alleles from both the maternal and paternal side. The pedigrees of both AM 78-3778 and AM 78-3704 display the same three sources of S. vernei (Supplementary data 6). The first is the breeding clone LGU 8 (developed by R.L. Plaisted, Cornell University, USA) using unnamed S. vernei material from Hans Ross (MPIZ, Germany). The second is clone V 24/20 which points back to Scottish work using CPC 2488-3 × CPC 2487-3. Third, VRN 1–3 represent material derived from S. vernei hybrid GLKS 58.1642-4 (Dellaert and Vinke 1987).
Cultivar INNOVATOR exhibits an exceptionally high level of resistance to the potato cyst nematode G. pallida, which is derived from progenitor AM 78-3778. Hence, we filtered on INNOVATOR as the resistant clone (50K SNPs), selected SNPs from chromosome 5 (5352 SNPs remain) with an allele count <20 (615 SNPs remain), and used ARRAN PILOT, BINTJE, CHARLOTTE, CIVA, DAISY, GOLDEN WONDER, HOME GUARD, KATAHDIN, ULTIMUS and YAM as negatives for G. pallida resistance. This resulted in 294 remaining SNPs according to the pattern shown in Fig. 6.

Graphical genotyping image of 83 tetraploid cultivars (columns) with 294 SNPs (in rows) from potato chromosome 5 where Gpa5 haploblock sharing and size of linkage drag is visualized. The reference panel of 83 cultivars (columns) has eight cultivars with resistance against cyst nematodes Globodera pallida (race pa2/3). The SNPs are located on superscaffolds (DMB numbers) as defined by PGSC (2011), and specific coordinates are given in suppl. material 6. The introgressed haplotypes of AVENANCE, INNOVATOR and WISENT comprise the entire north arm of chromosome 5. Cultivar FESTIEN has lost most linkage drag with an introgression segment of ~0.7 Mb
The graphical genotyping image shown in Fig. 6 shows a potentially erroneous orientation of DMB103. A reorientation of DMB103 would nullify the gap.
The transitions from green to red in the columns of Fig. 6 also display the positions of the historical recombination events, which caused a reduction of the linkage drag associated with Gpa5 introgression. The cultivars AVENANCE (2005), INNOVATOR (1999) and WISENT (2005) have the largest introgression segment, followed by NOMADE (1995). In MERCATOR (1999) and KARTEL (1994), the introgression segment is again shortened and linkage drag is minimal in FESTIEN (2000). The shortening of the introgression segment (which is the same as the removal of linkage drag) does not follow a trend from older to more recently released cultivars. Therefore, the historical recombination events must have taken place in between generations deeper in the pedigrees.
The shortest segment observed in cv. FESTIEN is consistent with the location of the HC marker (Achenbach et al. 2009). The HC marker is known to produce the least amount of false negatives of all markers developed to select Gpa5. To compare the physical coordinates of the HC marker relative to the graphical location of Gpa5 (Fig. 6), a BLAST search was performed on the pseudomolecules of PGSC v4.03, using the primer sequences of the HC marker. BLAST indeed finds the best hit within DMB424 at superscaffold coordinates 174417..174688.
Negative results
Unfortunately, we were unable to find SNPs indicating the Gro1–4 gene on chromosome 7 involved in resistance against Globodera rostochiensis nematodes (also known as the Fb gene, Paal et al. 2004). This effort failed because it is unknown how the experimental material, from which the gene was cloned, is related to cultivars that entered the market.
Discussion
Intuitively one may easily accept that application of a few selection criteria imposed on a data set with 129,156 SNPs allows the graphical visualization of haploblocks. To explain the underlying principles is, however, not as easy. SNPs are binary characters which allow one to separate haplotypes into two groups: a group with haplotypes that share the SNP allele and a group of haplotypes that share the alternative variant. The number of haplotypes in the potato gene pool is, however, much larger and can be deduced from the large number of SNPs with a low minor allele frequency (MAF) as shown in Fig. 2. The average MAF is 0.14 only. Moreover, 17.4% of all SNPs have a MAF <1%, and 39.1% have a MAF <5% (Uitdewilligen et al. 2013). Supposing that all SNPs with an MAF <5% are haplotype specific (hs-SNPs), this allows one to imagine that the gene pool is comprised of at least a dozen different haplotypes per locus. Apart from the hs-SNPs, the remaining SNPs with a higher MAF may consign haplotypes into two groups, without any reason to assume that the alleles within such a group have anything in common.
Graphical genotyping thus relies on the ability to distinguish specific minor alleles because of the presence of hs-SNPs against the many other haplotypes which share the absence of this minor hs-SNP allele. In other words, haplotype specificity refers to the minor allele. The major allele is not haplotype specific at all.
A negative result of graphical genotyping was the inability to find SNPs indicating the Gro1–4 locus. Possibly the Gro1–4 gene is hardly deployed in variety breeding and, therefore, escapes detection in this limited variety panel. Furthermore, Ro5-resistant cultivars descending from S. spegazzinii have an overly complex pedigree resulting in ambiguity if they share any most recent common ancestor. We also obtained negative results for several other single locus traits, such as flesh colour (Wolters et al. 2010), tuber shape (Van Eck et al. 1994) and maturity (Kloosterman et al. 2013). This is likely to be due to a violation of the assumption of identity-by-descent. If a phenotype is not the result of a single (recent) mutation, also indicated as genetic heterogeneity, then other (statistical) methods will also fail. In addition, the size of the haploblock may be too short to allow the identification of a meaningful tract of SNPs in a graphical genotyping image. Finally, it should be noted that although haplotypes are defined by SNPs, it is not necessarily that any of these SNPs are haplotype specific. We, therefore, conclude that the power of graphical genotyping and the power of statistical methods equally suffer from trait heterogeneity and genetic heterogeneity.
Another notion to understand this application of graphical genotyping in unrelated tetraploids is based on the breeding history of potato cultivars. During a century of potato breeding and approximately one meiosis per decade (Love 1999), large haploblocks can be expected. The gene pool has a limited number of founders and donors of resistance and, therefore, identity-by-descent is obvious for haplotypes shared by related cultivars. These aspects contribute to long haploblocks. Shorter haploblocks associated with traits can also appear by chance. The probability for such random effects is also influenced by the nucleotide diversity between haplotypes.
Finally, within a given level of LD, nucleotide diversity and founders, the number of SNPs needs to exceed a certain minimal number. Only with a large number of SNPs clear stripes will appear within the image, where the length of the stripes enables the visual identification of haplotype sharing. Future work may show that the number of markers comprised by each stripe is indicative of the statistical significance for the conclusion of haplotype sharing or IBD. In Table 2, a summary is given of the many aspects that contribute to the success or failure of graphical genotyping. Most aspects have a similar impact on the success of genome-wide association studies (GWAS).
We have mapped a locus involved in PVY resistance derived from CPC 2093. The putative resistance gene in the locus was tentatively named Ny (o,n)sto following guidelines (Valkonen et al. 1996), because the resistant parent (FESTIEN) of the gene mapping population expressed HR to both PVYO and PVYNTN. However, the mapping population was studied only for segregation of resistance to PVYNTN and remains to be tested with PVYO before involvement of the same locus in resistance to both PVY strains can be confirmed. Furthermore, we showed that FESTIEN responds with HR to PVA. Co-segregation of resistance to PVY and PVA is known in S. stoloniferum (Cockerham 1970), but co-segregation of the gene markers and HR to PVA should be tested to identify the PVA resistance locus.
In view of HR conferred by Ny (o,n)sto and ER conferred by Ry sto and Ry-f sto (Brigneti et al. 1997; Flis et al. 2005; Song et al. 2005; Valkonen et al. 2008), the genes underlying these resistances should be different. Intriguingly, the PVY-resistant variety TIVOLI, a first-generation descendant of the resistant parent I 1039, which was used by Brigneti et al. (1997), was positive for the SNP marker from this paper according to marker assays tested at HZPC (personal communication). A second discrepancy between marker data, pedigree information and resistance phenotype that is relevant in the context of possible shared identity with Ny (o,n)sto relates to the Ny-2 locus (Szajko et al. 2014). The Ny-2 locus mapped in the German variety ROMULA relased in 2002. ROMULA descends from VE 79-113, which descends via W 72-38-720 from Y 66-13-636). ROMULA was characterized to confer HR to PVY, whereas SANTÉ (an F1 from Y 66-13-636) showed ER. Both ROMULA and SANTÉ have identical marker scores and these should be identical by descend according to their pedigrees. Not only Szajko et al. (2014) but also Heldák et al. (2007) classified SANTÉ as ER based on low ELISA readings. When judging from the ROMULA perspective, the Ny (o,n)sto and Ny-2 locus could be identical, but this is at odds with the phenotypic classification of SANTÉ being ER. These discrepancies may be explained by two mechanisms. First, the genes (Ry) for ER to PVY are epistatic to the genes (Ny) for HR and, hence, only the resistance phenotype conferred by Ry can be observed in the presence of both types of genes in the same plant (Valkonen et al. 1994). Second, the role of Ry and Ny is to recognize the virus, which triggers the signalling cascades for effective defence responses. However, allelic variation in the genes involved in signalling for defence, or in the defence mechanism itself, may render the defence less effective, which results in an altered phenotypic outcome. For example, detectable accumulation of PVY is usually prevented in potato plants carrying Ry, no matter whether plants are sap-inoculated or graft-inoculated (Valkonen et al. 1994; Hämäläinen et al. 1997). Nevertheless, some potato genotypes carrying Ry may contain low but detectable PVY titres and systemic infection following graft inoculation, which results in small necrotic lesions and/or veinal necrosis in the top leaves. While Ny normally prevents movement of PVYO from the inoculated leaves to other parts of the plant, genetic variability in the defence signalling cascades may cause temperature sensitivity, which allows systemic infection at elevated temperatures (Valkonen 1997; Szajko et al. 2014). Wider testing and comparative analysis with, e.g. marker B11.6 (Szajko et al. 2014) is required.
Our SNP markers matching the haplotype shared by EOS, FESTIEN and Y 66-13-636 allow marker-assisted breeding. Subsequent validation with an Infinium SNP array on a wider panel of 537 cultivars (Vos et al. 2015) could identify PVY-resistant or -susceptible varieties without any discrepancy. We conclude that graphical genotyping is not only suitable to map loci in bi-parental mapping populations, but also in panels of distantly related cultivars. The graphical genotyping patterns observed here suggest the presence of specific haplotypes which are uniquely tagged by tracts of haplotype-specific SNPs.
Graphical genotyping was initially proposed as a tool in mapping studies, but it also makes singleton observations visible to allow correction of erroneous genotyping results. Whereas the data structure from a panel of cultivars (as shown in Fig. 1b) suggests that correction of genotyping errors or the identification of mislabelled of plant material is not straightforward, this paper shows that skilful use of graphical genotyping images in cultivar panels can assist in making various interpretations of the data, including observation of trait heterogeneity and genetic heterogeneity.
Author contribution statement
Conceived and designed the experiments: HvE. Performed the experiments: JU, HL, NdV, JV. Analysed the data: HvE, JU, PV, JV. Wrote the paper: HvE, PV, JV.
References
Achenbach U, Paulo J, Ilarionova E, Lübeck J, Strahwald J, Tacke E, Hofferbert H-R, Gebhardt C (2009) Using SNP markers to dissect linkage disequilibrium at a major quantitative trait locus for resistance to the potato cyst nematode Globodera pallida on potato chromosome V. Theor Appl Genet 118:619–629
Bakker E, Borm T, Prins P, van der Vossen E, Uenk G, Arens M, De Boer J, Van Eck HJ, Muskens M, Vossen J, Van der Linden G, Van Ham R, Klein-Lankhorst R, Visser RGF, Smant G, Bakker J, Goverse A (2011) A genome-wide genetic map of NB-LRR disease resistance loci in potato. Theor Appl Genet 123(3):493–508
Bergelson J, Roux F (2010) ) Towards identifying genes underlying ecologically relevant traits in Arabidopsis thaliana. Nat Rev Genet 11(12):867–879
Brigneti G, Garcia-Mas J, Baulcombe DC (1997) Molecular mapping of the Potato virus Y resistance gene Ry sto in potato. Theor Appl Genet 94:198–203
Celebi-Toprak F, Slack SA, Jahn MM (2002) A new gene, Ny tbr , for hypersensitivity to Potato virus Y from Solanum tuberosum maps to chromosome IV. Theor Appl Genet 104(4):669–674
Cockerham G (1970) Genetical studies on resistance to potato viruses X and Y. Heredity 25:309–348
D’hoop BB, Keizer PL, Paulo MJ, Visser RGF, van Eeuwijk FA, van Eck HJ (2014) Identification of agronomically important QTL in tetraploid potato cultivars using a marker–trait association analysis. Theor Appl Genet 127(3):731–748
De Jong H, Proudfoot KG, Murphy AM (2001) The germplasm release of F87084, a fertile, adapted clone with multiple disease resistances. Am J Potato Res 78:141–149
Dellaert LM, Vinke HJ (1987) Testing potatoes for resistance to Globodera pallida pathotype Pa-3; resistance spectra of plant genotypes and virulence spectra of Pa-3 isolates. Revue Nématol 10(4):445–453
Fenlon JS, Sopp PI (1991) Some statistical considerations in the determination of thresholds in ELISA. Ann Appl Biol 119:177–189
Finkers-Tomczak A, Danan S, van Dijk T, Beyene A, Bouwman L, Overmars H, van Eck HJ, Goverse A, Bakker J, Bakker E (2009) A high-resolution map of the Grp1 locus on chromosome V of potato harbouring broad-spectrum resistance to the cyst nematode species Globodera pallida and Globodera rostochiensis. Theor Appl Genet 119(1):165–173
Flis B, Hennig J, Strzelczyk-Żyta D, Gebhardt C, Marczewski W (2005) The Ry-f sto gene from Solanum stoloniferum for extreme resistant to Potato virus Y maps to potato chromosome XII and is diagnosed by PCR marker GP122718 in PVY resistant potato cultivars. Mol Breed 15(1):95–101
Hämäläinen JH, Watanabe KN, Valkonen JPT, Arihara A, Plaisted RL, Pehu E, Miller L, Slack SA (1997) Mapping and marker-assisted selection for a gene for extreme resistance to Potato virus Y. Theor Appl Genet 94(2):192–197
Hämäläinen JH, Sorri VA, Watanabe KN, Gebhardt C, Valkonen JPT (1998) Molecular examination of a chromosome region that controls resistance to potato Y and A potyviruses in potato. Theor Appl Genet 96(8):1036–1043
Hehl R, Faurie E, Hesselbach J, Salamini F, Whitham S, Baker B, Gebhardt C (1999) TMV resistance gene N homologues are linked to Synchytrium endobioticum resistance in potato. Theor Appl Genet 98(3–4):379–386
Heldák J, Bežo M, Štefúnová V, Galliková A (2007) Selection of DNA markers for detection of extreme resistance to Potato virus Y in tetraploid potato (Solanum tuberosum L.) F1 progenies. Czech J Genet Plant Breed 43(4):125–134
Hosaka K, Hosaka Y, Mori M, Maida T, Matsunaga H (2001) Detection of a simplex RAPD marker linked toresistance to potato virus Y in a tetraploid potato. Am J Potato Res 78(3):191–196
Jupe F, Pritchard L, Etherington GJ, MacKenzie K, Cock PJ, Wright F, Sharma SK, Bolser D, Bryan GJ, Jones JDG, Hein I (2012) Identification and localisation of the NB-LRR gene family within the potato genome. BMC Genom 13(1):75
Kloosterman B, Abelenda JA, del Mar Carretero Gomez M, Oortwijn M, de Boer JM, Kowitwanich K, Horvath BM, van Eck HJ, Smaczniak C, Prat S, Visser RGF, Bachem CWB (2013) Naturally occurring allele diversity allows potato cultivation in northern latitudes. Nature 495(7440):246–250
Love SL (1999) Founding clones, major contributing ancestors, and exotic progenitors of prominent North American potato cultivars. Am J Potato Res 76(5):263–272
Milne I, Shaw P, Stephen G, Bayer M, Cardle L, Thomas WTB, Flavell AJ, Marshall D (2010) Flapjack—graphical genotype visualization. Bioinformatics 26(24):3133–3134
Motte H, Vercauteren A, Depuydt S, Landschoot S, Geelen D, Werbrouck S, Goormachtig S, Vuylsteke M, Vereecke D (2014) Combining linkage and association mapping identifies RECEPTOR-LIKE PROTEIN KINASE1 as an essential Arabidopsis shoot regeneration gene. Proc Natl Acad Sci USA 111(22):8305–8310
Moury B, Caromel B, Johansen E, Simon V, Chauvin L, Jacquot E, Kerlan C, Lefebvre V (2011) The helper component proteinase cistron of Potato virus Y induces hypersensitivity and resistance in potato genotypes carrying dominant resistance genes on chromosome IV. Mol Plant-Microbe Interact 24(7):787–797
Nie X, Liang Z, Nie B, Murphy A, Singh M (2015) Studies on varietal response to different strains of Potato virus Y (PVY) reveal hypersensitive resistance in exploits to PVYO and extreme resistance in F87084 to all tested strains. Am J Potato Res 92(1):23–31
Paal J, Henselewski H, Muth J, Meksem K, Menéndez CM, Salamini F, Ballvora A, Gebhardt C (2004) Molecular cloning of the potato Gro1–4 gene conferring resistance to pathotype Ro1 of the root cyst nematode Globodera rostochiensis, based on a candidate gene approach. Plant J 38(2):285–297
PGSC—Potato Genome Sequencing Consortium (2011) Genome sequence and analysis of the tuber crop potato. Nature 475(7355):189–195
Rajamäki M, Merits A, Rabenstein F, Andrejeva J, Paulin L, Kekarainen T, Kreuze JF, Forster RLS, Valkonen JPT (1998) Biological, serological and molecular differences among isolates of potato A potyvirus. Phytopathology 88:311–321
Ross H (1986) Potato breeding—problems and perspectives. J Plant Breed Suppl 13
Rouppe van der Voort J, Van der Vossen E, Bakker E, Overmars H, Van Zandvoort P, Hutten R, Klein Lankhorst R, Bakker J (2000) Two additive QTLs conferring broad-spectrum resistance in potato to Globodera pallida are localized on resistance gene clusters. Theor Appl Genet 101(7):1122–1130
Sato M, Nishikawa K, Komura K, Hosaka K (2006) Potato virus Y resistance gene, Ry chc , mapped to the distal end of potato chromosome 9. Euphytica 149:367–372
Sharma SK, Bolser D, de Boer J, Sønderkær M, Amoros W, Carboni MF, Amoros de la Cruz G, Di Genova A, Douches DS, Eguiluz M, Guo X, Guzman F, Hackett CA, Hamilton JP, Li G, Li Y, Lozano R, Maass A, Marshall D, Martinez D, McLean K, Mejía N, Milne L, Munive S, Nagy I, Ponce O, Ramirez M, Simon R, Thomson SJ, Torres Y, Waugh R, Zhang Z, Huang S, Visser RGF, Bachem CWB, Sagredo B, Feingold SE, Orjeda G, Veilleux RE, Bonierbale M, Jacobs JME, Milbourne D, Martin DMA, Bryan GJ (2013) Construction of reference chromosome-scale pseudomolecules for potato: integrating the potato genome with genetic and physical maps. G3: Genes| Genomes| Genet 3(11):2031–2047
Song Y-S, Schwarzfischer A (2008) Development of STS markers for selection of extreme resistance (Ry sto ) to PVY and maternal pedigree analysis of extremely resistant cultivars. Am J Potato Res 85:159–170
Song Y-S, Hepting L, Schweizer G, Hartl L, Wenzel G, Schwarzfischer A (2005) Mapping of extreme resistance to PVY (Ry sto ) on chromosome XII using anther-culture-derived primary dihaploid potato lines. Theor Appl Genet 111:879–887
Szajko K, Chrzanowska M, Witek K, Strzelczyk-Żyta D, Zagórska H, Gebhardt C, Hennig J, Marczewski W (2008) The novel gene Ny-1 on potato chromosome IX confers hypersensitive resistance to Potato virus Y and is an alternative to Ry genes in potato breeding for PVY resistance. Theor Appl Genet 116:297–303
Szajko K, Strzelczyk-Żyta D, Marczewski W (2014) Ny-1 and Ny-2 genes conferring hypersensitive response to Potato virus Y (PVY) in cultivated potatoes: mapping and marker-assisted selection validation for PVY resistance in potato breeding. Mol Breed 34(1):267–271
Tian YP, Valkonen JPT (2013) Genetic determinants of Potato virus Y required to overcome or trigger hypersensitive resistance to PVY strain group O controlled by the gene Ny in potato. Mol Plant-Microbe Interact 26:297–305
Tian YP, Hepojoki J, Ranki H, Lankinen H, Valkonen JPT (2014) Analysis of Potato virus Y coat protein epitopes recognized by three commercially available monoclonal antibodies. PLoS One 9:e115766
Tomczyńska I, Jupe F, Hein I, Marczewski W, Śliwka J (2014) Hypersensitive response to Potato virus Y in potato cultivar Sárpo Mira is conferred by the Ny-Smira gene located on the long arm of chromosome IX. Mol Breed 34(2):471–480
Uitdewilligen JGAML, Wolters AMA, D’hoop BB, Borm TJA, Visser RGF, Van Eck HJ (2013) A next-generation sequencing method for genotyping-by-sequencing of highly heterozygous autotetraploid potato. PloS One 8(5):e62355
Valkonen JPT (1997) Novel resistances to potyviruses in tuber-bearing potato species, and temperature-sensitive expression of hypersensitive resistance to potato virus Y. Ann Appl Biol 130:91–104
Valkonen JPT (2007) Chapter 28—viruses: economical losses and biotechnological potential. In: Vreugdenhil D, Bradshaw J, Gebhardt C, Govers F, Mackerron D, Taylor M, Ross H (eds) Potato biology and biotechnology. Elsevier Science B.V., Amsterdam, pp 619–641
Valkonen JPT, Slack SA, Plaisted RL, Watanabe K (1994) Extreme resistance is epistatic to hypersensitive resistance to potato virus Yo in a Solanum tuberosum ssp. andigena-derived potato genotype. Plant Dis 78:1177–1180
Valkonen JPT, Jones RAC, Slack SA, Watanabe KN (1996) Resistance specificities to viruses in potato: standardization of nomenclature. Plant Breed 115:433–438
Valkonen JPT, Wiegmann K, Hämäläinen JH, Marczewski W, Watanabe KN (2008) Evidence forutility of the same PCR-based markers for selection of extreme resistance to Potato virus Y controlled by Rysto of Solanum stoloniferum derived from different sources. Ann Appl Biol 152(1):121–130
Van Berloo R (1999) GGT: software for the display of graphical genotypes. J Hered 90(2):328–329
Van Berloo R (2008) GGT 2.0: versatile software for visualization and analysis of genetic data. J Hered 99(2):232–236
Van Berloo R, Hutten RCB, Van Eck HJ, Visser RGF (2007) An online potato pedigree database resource. Potato Res 50(1):45–57
Van Eck HJ, Jacobs JM, Stam P, Ton J, Stiekema WJ, Jacobsen E (1994) Multiple alleles for tuber shape in diploid potato detected by qualitative and quantitative genetic analysis using RFLPs. Genetics 137(1):303–309
Van Os H, Stam P, Visser RGF, Van Eck HJ (2005) SMOOTH: a statistical method for successful removal of genotyping errors from high-density genetic linkage data. Theor Appl Genet 112(1):187–194
Vos PG, Uitdewilligen JG, Voorrips RE, Visser RGF, Van Eck HJ (2015) Development and analysis of a 20K SNP array for potato (Solanum tuberosum): an insight into the breeding history. Theor Appl Genet 128(12):2387–2401
Wolters A-MA, Uitdewilligen JGAML, Kloosterman BA, Hutten RCB, Visser RGF, van Eck HJ (2010) Identification of alleles of carotenoid pathway genes important for zeaxanthin accumulation in potato tubers. Plant Mol Biol 73(6):659–671
Young ND, Tanksley SD (1989) Restriction fragment length polymorphism maps and the concept of graphical genotypes. Theor Appl Genet 77(1):95–101
Yun SJ, Gyenis L, Bossolini E, Hayes PM, Matus I, Smith KP, Steffenson BJ, Tuberosa R, Muehlbauer GJ (2006) Validation of quantitative trait loci for multiple disease resistance in barley using advanced backcross lines developed with a wild barley. Crop Sci 46(3):1179–1186
Acknowledgements
This research was partially supported by a grant from the Dutch technology foundation STW, project WPB-7926. Peter Vos is supported by a grant from potato breeding companies Agrico Research B.V, Averis Seeds B.V, HZPC B.V, KWS POTATO B.V. and Meijer B.V. We thank Peter Bourke for critical reading of the manuscript.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors are not aware of any affiliations, memberships, funding, or financial holdings that might be perceived as affecting the objectivity of this publication.
Additional information
Communicated by J. Wang.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
van Eck, H.J., Vos, P.G., Valkonen, J.P.T. et al. Graphical genotyping as a method to map Ny (o,n)sto and Gpa5 using a reference panel of tetraploid potato cultivars. Theor Appl Genet 130, 515–528 (2017). https://doi.org/10.1007/s00122-016-2831-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00122-016-2831-y