
Abstract
The extreme acid conditions required for scorodite (FeAsO4·2H2O) biomineralization (pH below 1.3) are suboptimal for growth of most thermoacidophilic Archaea. With the objective to develop a continuous process suitable for biomineral production, this research focuses on growth kinetics of thermoacidophilic Archaea at low pH conditions. Ferrous iron oxidation rates were determined in batch-cultures at pH 1.3 and a temperature of 75°C for Acidianus sulfidivorans, Metallosphaera prunea and a mixed Sulfolobus culture. Ferrous iron and CO2 in air were added as sole energy and carbon source. The highest growth rate (0.066 h−1) was found with the mixed Sulfolobus culture. Therefore, this culture was selected for further experiments. Growth was not stimulated by increase of the CO2 concentration or by addition of sulphur as an additional energy source. In a CSTR operated at the suboptimal pH of 1.1, the maximum specific growth rate of the mixed culture was 0.022 h−1, with ferrous iron oxidation rates of 1.5 g L−1 d−1. Compared to pH 1.3, growth rates were strongly reduced but the ferrous iron oxidation rate remained unaffected. Influent ferrous iron concentrations above 6 g L−1 caused instability of Fe2+ oxidation, probably due to product (Fe3+) inhibition. Ferric-containing, nano-sized precipitates of K-jarosite were found on the cell surface. Continuous cultivation stimulated the formation of an exopolysaccharide-like substance. This indicates that biofilm formation may provide a means of biomass retention. Our findings showed that stable continuous cultivation of a mixed iron-oxidizing culture is feasible at the extreme conditions required for continuous biomineral formation.
Similar content being viewed by others

Introduction
Biomineral production has the potential to become an active field of research for the development of new materials and applications. Some of these applications require the adaptation of microorganisms to extreme conditions. This is the case for the biogenic production of scorodite by thermoacidophilic microorganisms, where temperatures above 60°C and a pH below 1.3 are needed to favour the formation of highly crystalline materials (Gonzalez-Contreras et al. 2010). Scorodite biomineral formation needs less than 1 g L−1 of Fe (Gonzalez-Contreras et al. 2009). This implies that these thermoacidophilic microorganisms must sustain growth below their optimal substrate concentrations and pH.
Although previous efforts to produce biominerals using thermoacidophilic microorganisms were successful, more information about the involved microorganisms is needed for the application in a continuous process. Several studies in engineered bioleaching operations describe how Sulfolobales Archaea mediate the oxidation of mineral sulphides through simultaneous sulphur and iron oxidation (Norris et al. 2000; Norris 2006; Dinkla et al. 2009; Etzel et al. 2008; Mikkelsen et al. 2007). Thermophile conditions (60–80°C) in a tank-bioleaching reactor are required when a significant portion of the copper is present as refractory primary copper sulphides such as chalcopyrite (Plessis et al. 2006). Growth parameters reported for Sulfolobales species are summarized in Table 1, which includes the dominant genera of Sulfolobus, Metallosphaera and Acidianus (Huber and Stetter 1998; Norris 2006; Ralings 2006). The low growth rate and low cell yield of thermoacidophilic cultures are known bottlenecks for their application in continuous bioreactors (Bertoldo et al. 2004) and for bioleaching of sulphide minerals (Huber and Stetter 1998; Nemati and Harrison 2000). In those applications, Sulfolobales species oxidized high ferrous iron concentrations of above 10 g L−1 Fe, at the optimum range pH for growth, pH 2–2.5. The iron oxidation rates for thermoacidophiles are about ten times as low as mesophiles iron-oxidizing acidophiles (Nemati and Harrison 2000). Information concerning thermoacidophilic microorganisms growing at suboptimal conditions and only using a single energy source, such as ferrous iron, is only scarcely available.
The objectives covered in this research are: (1) to select a suitable thermoacidophilic culture for ferrous iron oxidation at extremely low pH of 1.1–1.3, for later use in biomineralization reactors; (2) to determine how the addition of co-substrates (sulphur, carbon dioxide enrichment) affects the microorganisms growing at suboptimal growth conditions, i.e. low pH and low ferrous iron concentration; (3) to determine the kinetics of biological oxidation of ferrous iron in batch tests and in continuous reactors and (4) to evaluate the stability of continuous reactors operated at suboptimal conditions during a period of 1 year.
Materials and methods
Microorganism and medium
The Archaea Acidianus sulfidivorans (DSM 18786) and Metallosphaera prunae (DSM 10039) were obtained from the German Collection of Microorganism and Cell Cultures (DSMZ, Braunschweig, Germany). A. sulfidivorans originates from a sulphur-rich acidic edge of a hot spring (Plumb et al. 2007). M. prunae originates from hot deposits in a slag heap of a uranium mine (Fuchs et al. 1996). Selected physiological characteristics of A. sulfidivorans and M. prunae are shown in Table 1. The mixed culture, kindly provided by M. Gericke from Mintek (Randburg, South Africa), was collected from a hot, sulphur-rich coal dump (Gericke and Pinches 1999). The mixed culture has a reported optimum growth temperature of 70°C and a pH optimum of 1.8 (Gericke and Pinches 1999). The dominant archaeon in this culture was Acidianus brierleyi (98–99%). Metallosphaera sedula (1–2%) and very small numbers of Sulfolobus spp. were also present in the culture (Dinkla et al. 2009).
The defined growth medium used for the thermophilic acidophiles contained: 1 g L−1 (NH4)2SO4, 0.1 g L−1 KCl, 0.5 g L−1 MgSO4·7H2O and 0.5 g L−1 K2HPO4 as described by Plumb et al. (2002). Micronutrients were added according to Gonzalez et al. (2010). All chemicals used were analytical-reagent grade. Ferrous iron stock solutions were prepared by dissolving iron (II) sulphate heptahydrate (FeSO4·7H2O) at several concentrations in 0.1 mol L−1 sulphuric acid (H2SO4). Biologically produced elemental sulphur (biosulphur) was used as sulphur source. Biosulphur was obtained from a full-scale biogas treatment facility (Eerbeek, the Netherlands) in which H2S is biologically oxidized to elemental sulphur. Biosulphur contains approximately 5% organic matter (Janssen et al. 2009). Biosulphur particles are hydrophilic and therefore easily dispersible in water, as opposed to chemically produced sulphur (Janssen et al. 2009).
Experimental procedures
Three series of batch experiments and two continuous stirred tank reactors (CSTR) experiments were carried out.
Batch tests
Serum bottles of 100 mL, closed with a butyl rubber stopper and a crimped aluminium seal were used as incubation vessels. One series of bottles was used for liquid sampling and another for gas sampling. The headspace/liquid volumetric ratio in the bottles was such that a 50% excess of oxygen (in air) was present relative to the stoichiometric amount needed for complete ferrous iron oxidation. The bottles were inoculated 10% v/v with the primary culture and incubated in a shaking incubator at 150 rpm and a temperature of 75°C. Ferrous iron oxidation was monitored by measuring ferrous iron depletion in the medium and oxygen depletion in the headspace. Liquid samples were taken with a syringe, filtered over a 0.45-μm cellulose acetate membrane filter (Spartan 30, Whatman, Germany) and analysed for pH, redox, cell count and ferrous and ferric iron concentration.
Batch experiment 1. Selection of thermoacidophilic archaeal culture
For A. sulfidivorans, M. prunae and the mixed culture, ferrous iron oxidation rates were determined at 1 g L−1. Bottles were incubated at 75°C. The mixed culture was also cultivated at 70°C [reported optimum temperature (Dinkla et al. 2009)].
Batch experiment 2. Determination of ferrous iron oxidation rate by a thermoacidophilic mixed culture at 1 g L−1 ferrous iron
The growth of a mixed culture oxidizing ferrous iron at 75°C was evaluated at various conditions of pH, the presence of sulphur, carbon dioxide enrichment and ferrous and ferric concentrations. All bottles were incubated with 1 g L−1 ferrous (Fe2+), 5% v/v carbon dioxide enrichment and 100 mg L−1 of elemental sulphur as additional energy sources and the initial pH was adjusted to 1.7. These conditions were kept the same for all bottles, except when a specific parameter was the subject of study.
Batch experiment 3. Kinetics of ferrous iron oxidation by the thermoacidophilic mixed culture at 75°C
The ferrous iron kinetic experiments were carried out at pH 0.9 and 1.3. The ferrous iron oxidation rate was assessed at both pH conditions in a ferrous iron range between 0 to 11 g L−1. Specific growth rate and cell yield on ferrous iron were determined as well.
CSTR reactors
Two aerated CSTR were used, each with 1 L of working volume. The temperature was controlled at 75 ± 2°C. The reactors were equipped with electrodes for the measurement of oxygen, pH, redox potential and temperature. The air flow to the reactors was controlled by mass flow controllers at a rate of 0.4 volume of air per volume of medium per minute, using a stainless diffuser. The effluent air was dehumidified with condensers and the condensate was returned to the CSTR. No evaporation was observed.
Methods
Iron species in the aqueous phase were measured using Dr. Lange cuvette test LCK 320 for Fe2+ and ferric iron and a Xion 500 spectrophotometer (Hach-Lange, Germany). Redox potential and pH were measured with glass electrodes QR480X-Pt billed-S8 and QP181X-S8 (Prosense, The Netherlands), respectively. Slope calibration of pH electrode was made with pH 1 and pH 4 buffers. Dissolved oxygen was measured using an InPro 6800 oxygen sensor (Mettler Toledo, Germany) connected to a Stratos e2402-oxy meter (Knick, Germany). The oxygen sensor was calibrated at 0% with nitrogen and at 100% with water-saturated air. Cells were counted in a Bürker-Türk chamber (Brand, Germany). In addition, biomass concentration was measured as the difference between the amount of total nitrogen and the total nitrogen in centrifuged samples (15 min, 12,000 rpm). The method is based on the absorbance of nitrophenol with the Dr. Lange cuvette test LCK238 (Hach-Lange, Germany).
Carbon dioxide and oxygen in the gas headspace of the bottles were determined with a gas chromatograph (Fisons Instruments GC 8000) equipped with two columns: 1.5 m × 1/8 in. teflon packed with Chromosorb 108 (60–80 mesh) and 1.2 m × 1/8 in. stainless steel packed with molecular sieve 5A (60–80 mesh). Both columns were connected in parallel with a split of 1:1. The carrier gas was helium with a flow rate of 45 mL min−1. Temperatures were 60°C for the column, 110°C for the injection port and 100°C for the thermal conductivity detector.
Autotrophic growth through elemental sulphur oxidation to sulphate was calculated by the difference between the oxygen consumed by ferrous iron oxidation and the consumption of total oxygen in the headspace of the bottles. The sulphate concentration was not measured as the concentration of sulphate in the medium (0.1 mol L−1 sulphuric acid medium) was too high to detect a small increase due to sulphur oxidation.
The morphology of the culture was investigated with scanning electron microscopy (SEM). Samples were prepared by mild centrifugation, after which pellets were fixed in glutaraldehyde 2.5% and dehydrated with a grades series of ethanol–water mixtures (70% to 100%). Then, the samples were dried by the critical point method and glued on a copper sample holder using conductive carbon tape and silver tape and sputter coated with 20 nm platinum (JFC 1200, JEOL, Japan). The surfaces were analysed with a FESEM (JEOL 6300 F, Tokyo, Japan) at room temperature, a working distance of 15 mm and SE detection at 3.5 kV. All images were recorded digitally (Orion, 6 ELI sprl., Belgium) at a scan rate of 100 s (full frame) at a size of 2,528 × 2,030, 8 bit. Noise reduction and resizing of the images were done with Adobe Photoshop CS. Besides, SEM–energy dispersive X-ray spectroscopy (INCA energy, Oxford Instruments Analytical, High Wycombe, England) was used to measure several elements in the samples (C, N, O and S).
Precipitates from the reactor were collected by filtration using paper filters (Schleicher & Schuell 589/1, pore size 12–25 μm), washed with 0.1 M sulphuric acid and dried at 60°C. X-ray diffraction (XRD) was used to identify the nature of the solid materials. Wide angle X-ray scattering powder diffractograms were recorded on a Philips PC-APD diffractometer in the reflection geometry in the angular range 5–50° (2θ), with a step size of 0.02° (2θ) and an acquisition time of 0.6 s per step. The CuKα1 radiation from the anode, generated at 40 kV and 50 mA, was monochromatized using a 15-μm Ni foil (λ = 0.1542 nm). The diffractometer was equipped with a 1° divergence slit, a 0.2-mm receiving slit and a 1° scatter slit.
Results
Batch experiment 1: selection of thermoacidophilic archaeal culture at 75°C, oxidizing 1 g L−1 of ferrous iron
The mixed culture and M. prunae had a higher ferrous iron oxidation rate (0.31 g L−1 d−1) compared to A. sulfidivorans (0.01 g L−1 d−1) (data not shown). Furthermore, the mixed culture had a higher ferrous iron oxidation rate at 75°C (0.5 g L−1 d−1) than at the reported optimum temperature of 70°C (0.34 g L−1 d−1) (data not shown). In previous experiments with A. sulfidivorans at the same conditions, we observed a higher ferrous iron oxidation rate of 0.04 g L−1 d−1 (Gonzalez-Contreras et al. 2010) than in the experiments described here. The lower ferrous iron oxidation rate observed in the current experiments was probably because A. sulfidivorans needs an additional energy source such as sulphur or mineral sulphide (Plumb et al. 2007). The mixed culture was selected for further experiments because of its high ferrous iron oxidation rate at 75°C in comparison with the pure cultures.
Batch experiment 2: determination of ferrous iron oxidation rate by a thermoacidophilic mixed culture at 75°C and 1 g L−1 of ferrous iron
A summary of the results is shown in Fig. 1.

Influence of initial pH, carbon dioxide enrichment in air, ferric iron concentration and sulphur concentration on the oxidation rate of 1 g L−1 ferrous iron (lines) and on the cell growth (bars). The experiments were carried out at 75°C using the mixed thermoacidophilic culture. Error bars of the duplicates are displayed in the plots. The bottles were inoculated with 3.2 107 cells mL−1
pH effect
The favourable pH range for the mixed culture was pH 1.4 to 2. At pH 1.2, a small drop of cell concentration was measured even though ferrous iron was completely oxidized. The microorganisms were inactive at pH 0.8.
Carbon dioxide enrichment in air
Air enriched with carbon dioxide up to 1.4% resulted in a small increase in the ferrous iron oxidation rate. However, it was observed that the ferrous iron oxidation rate decreased when carbon dioxide was increased up to 5%. The cell count did not substantially increase with respect to carbon dioxide addition. This was confirmed by the constant total biomass–nitrogen concentration of 1.5 mg N L−1. These results imply that the microorganisms were not limited by carbon dioxide in the gas phase.
The effect of sulphur as a secondary energy source
In bottles supplied with 0.5 and 1 g L−1 of elemental sulphur, a pH drop from 1.63 to 1.54 was measured due to proton formation from sulphur oxidation (2S° + 3O2 + 2H2O → 2H2SO4) (Bosecker 1997). Sulphur oxidation in the bottles was monitored by oxygen consumption. When oxygen depletion was only the result of oxidation of 0.96 to 0.99 mmol Fe2+ per bottle (~1 g L−1 Fe2+), a stoichiometric oxygen consumption of 0.22 to 0.24 mmol O2 per bottle was found. When sulphur was added, the total oxygen consumption increased to the values of 0.33 to 0.48 mmol O2 per bottle, i.e. oxygen was consumed by the microorganisms to oxidize sulphur. The supply of sulphur as an extra energy source did not increase the carbon dioxide consumption of 0.024 mmol or the number of active cells of 3.2 107 cell mL−1 during the experiment.
The effect of ferric iron concentration
The addition of 5 or 10 g L−1 ferric iron resulted in a drop of the pH from 1.5 to 0.8, probably due to the precipitation of ferric oxyhydroxide which is an acid-generating reaction. As a consequence of the pH drop, the cells were inactive and a ferrous iron conversion of only 10% was observed.
Batch experiment 3: batch kinetics of ferrous iron oxidation by a thermoacidophilic mixed culture at 75°C
Two pH values were selected for the batch kinetics: a pH of 1.3 because this is the maximum for scorodite crystallization and a pH of 0.9 because this is the minimum pH for growth. Both values are suitable for scorodite biomineralization, although the lower pH is preferable because it will minimize the formation of other iron precipitates (Gonzalez-Contreras et al. 2010). Figure 2 shows that higher ferrous iron oxidation rates were obtained at a pH of 1.3 ± 0.07 than at a pH 0.9 ± 0.03. The specific growth rate and growth yield were calculated from the amount of ferrous iron oxidized by the cells and the increase of biomass measured as number of cells per millilitre. The growth–substrate relationship is shown in Fig. 3. The specific growth rate of cells in suspension was a function of limiting substrate concentration (ferrous iron) and pH (0.9 and 1.3). At ferrous iron concentrations below 1 g L−1, linear growth was observed. This indicates that the substrate concentration was much lower than the saturation concentration according to Monod kinetics. At higher ferrous iron concentrations, exponential growth was observed. Despite the lower specific growth rate at inhibiting ferrous iron concentrations (above 8 g L−1), cells were active as shown by the trend line in Fig. 2. The maximum specific growth rate and saturation constant were obtained by the reciprocal plot of the substrate versus the specific growth rate. The saturation constant was 0.18 g L−1 and the maximum specific growth rate was 0.066 h−1 at pH 1.3. The strong effect of the pH on the specific growth rate is shown in Fig. 3a. Figure 3b shows that at ferrous iron concentrations of 1 g L−1, the overall growth yield (observed) was higher (3 107 cells kg−1 ferrous iron) than at higher concentrations of 6 g L−1 (1 107 cells kg−1 ferrous iron).

Biological ferrous iron oxidation rate at different initial ferrous iron concentrations. The biological ferrous iron oxidation rate was evaluated at two pH values: filled squares pH = 1.3, filled triangles pH = 0.9. Error bars of the duplicates are displayed in the plots

Specific growth rate and yield of the mixed culture calculated at different initial ferrous iron concentrations. The kinetic parameters were evaluated at two pH values: filled squares pH = 1.3, filled triangles pH = 0.9. Error bars of the duplicates are displayed in the plots
Continuous bioreactor tests
The conditions selected for the start-up of the continuous reactors were based on the results from batch experiments. In two reactors (CSTR-1 and CSTR-2), the influence of dilution rate and influent ferrous iron concentration on the ferrous iron oxidation rate was determined. CSTR-1 was fed with 3 g L−1 and CSTR-2 with 6 g L−1 ferrous iron. The start-up strategy of the bioreactors was to increase the dilution rate (D) up to the point where D approaches the maximum specific growth rate (μ max). In both reactors, ferrous iron oxidation rates were determined at the different dilution rates.
The CSTR reactors were started at pH 2 and their adaptation to pH 1.1 was successfully achieved at a dilution rate of 0.01 h−1 in CSTR-1 and 0.02 h−1 in CSTR-2. The ferrous iron was almost completely (>90%) oxidized to ferric iron. This was reflected by the high redox potential of 500–600 mV. During periods when ferrous iron oxidation was incomplete, redox values were lower at 400–500 mV (Supplementary Fig. S1). Throughout the continuous experiments, the oxygen level in both reactors remained at 3.23 mg L−1, which is 87% of saturation at 75°C, showing that oxygen supply was not limiting growth.
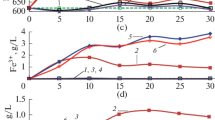
In CSTR-1, the increase of the dilution rate from 0.01 to 0.022 h−1 accelerated the ferrous iron oxidation rate from 0.4 to 1.4 g L−1 d−1. Further increase of the dilution rate after 190 days to a maximum of 0.036 h−1 resulted in an increase of the effluent ferrous iron concentration and a corresponding lower redox potential (Fig. S1-B), indicating washout of the cells (Fig. 4a). Subsequently, the reactor was again operated at lower dilution rates and stable iron oxidation was restored. The maximum specific growth rates were calculated using a Lineweaver–Burk plot. Specific growth rates of 0.022 and 0.037 h−1 were obtained at pH values of 1.1 and 1.2, respectively. The maximum specific growth rate at pH 1.3, as determined in batch culture, was 0.066 h−1.

Ferrous iron oxidation rates at different specific growth rates in (a) CSTR-1 and (b) CSTR-2. Legend: filled circles ferrous iron oxidation rate, empty squares dilution rate
In CSTR-2, the increase of ferrous iron supply in the influent from 3 to 6 g L−1 led to instability of the oxidation process (day 77–day 203). The ferrous iron oxidation rate fluctuated between 0.5 and 2.5 g L−1 d−1 and during the period that the reactor was supplied with 6 g L−1, the culture did not reach steady-state iron oxidation (Fig. 4b). Once the concentration was halved to 3 g L−1, the culture returned to steady-state ferrous iron oxidation with a rate of 1.5 g L−1 d−1 at pH 1.1 at a dilution rate of 0.02 h−1. The unsteady situation observed at 6 g L−1, ferrous iron in the influent suggests that elevated ferric iron concentrations may be inhibiting for the cells.
The high ferrous iron oxidation rate of 1.5 g L−1 d−1 was maintained in CSTR-2 during 80 days of stable operation, demonstrating the stability of the culture. Although high iron oxidation rates were obtained at the suboptimal pH of 1.1, the maximum specific growth rate was only one third of the maximum value of 0.066 h−1, achieved in the batch tests.
Morphology of the thermoacidophilic mixed culture
Based on cell morphology, the thermoacidophilic mixed culture can be classified as Sulfolobales-like. Figure 5a shows the lobed coccoid morphology. The size of the cells ranged from 0.5 to 1 μm. These results correspond to the reported morphology of the known Sulfolobales genera Methallosphaera, Sulfolobus and Acidianus (Huber and Stetter 1998).

a SEM photograph of the mixed culture growing in batch tests in ferrous iron at 75°C and pH of 1.5. Photograph with SE detection at 3 kV and magnification of ×25,000. b SEM photograph of the mixed culture growing in continuous reactors using ferrous iron at 75°C and pH of 1.2. EPS-like material was observed between cells and providing attachment to precipitates. Photograph with SE detection at 3 kV and magnification of ×35,000
In samples from the batch tests, nano-sized precipitates were observed on the cell surface (Fig. 5a). This indicates that the cell activity or the cell surface mediates the formation of these precipitates. The nano precipitates were easily separated and settled as a yellow powder, identified as K-jarosite by XRD analysis (Supplementary Fig. S2). Also, in continuous bioreactor tests, the microorganisms induced jarosite precipitation. During these continuous bioreactors tests, also the production of exopolysaccharide (EPS) like substances was observed (Fig. 5b and Supplementary Fig. S3). The EPS, which had a typical mucous appearance, was observed between cells and seemed to provide attachment sites for precipitates. The cell surface composition was randomly determined by SEM–EDX at 26% C, 25% Fe, 4% N, 9% S and 35% O.
Discussion
Selection of microorganisms and factors affecting the kinetics of iron oxidation by thermoacidophilic culture at 75°C and pH 1.1–1.3
From batch tests, a mixed culture was selected for further experiments because of its high ferrous iron oxidation rate at 75°C in comparison with the pure cultures. Several factors were studied on the biological oxidation of low ferrous iron concentrations needed for the scorodite process of 1 g L−1.
Supply of another substrate source or enrichment of carbon dioxide did not improve the cells’ growth but a high concentration of carbon dioxide resulted in a small increase of the iron oxidation rate. The results suggest that the cells may be sensitive to high carbon dioxide concentrations, which decrease the activity of the cells. To our best knowledge, carbon dioxide inhibition of thermoacidophilic microorganism has not been yet described. In contrast, for meso-acidophilic microorganisms, carbon dioxide inhibition has been described, although there is no general consensus about the limiting or inhibitory levels (Gericke et al. 2001; Witne and Phillips 2001; De Kock et al. 2004; Astudillo and Acevedo 2009).
A decrease of pH below 0.8 led to inactive microorganisms. The drastic effect of the pH is a bottleneck for the application of these microorganisms at extremely low pH process conditions. This suggests that a pH decrease in thermoacidophilic cultures must be taken into consideration in the bioleaching of sulphide minerals. The maximum specific growth rates obtained in the current research, of 0.066 h−1 at pH 1.3, are within the typical values reported in literature for thermoacidophilic microorganisms, even though we worked at lower, suboptimal pH conditions (Table 1).
Potential applications of the archaeal culture for biomineral formation
In the current study, we determined the kinetics of ferrous iron oxidation by a mixed thermoacidophilic culture in batch and continuous bioreactors at pH values and substrate concentrations below the optimum values for growth. In a continuous CSTR reactor, ferrous iron oxidation was obtained at pH of 1.1 and 75°C. The reactor had a steady ferrous iron oxidation rate of 1.5 g L−1 d−1 during 80 days at a dilution rate of 0.02 h−1, demonstrating the stability of the process during long-term operation.
The presence of polysaccharide-like material suggests that biofilm formation may be possible. This would facilitate biomass retention in suspended growth bioreactors. The formation of EPS by Archaea is scarcely reported for thermophiles (above 60°C) (Auernik et al. 2008) and also not for moderate thermophiles (45°C) (Becker et al. 2011; Zeng et al. 2011).
The apparent formation of EPS was stimulated during the continuous operations at a low pH value of 1.2 and not during the batch tests. This suggests that EPS could play a major role in shielding the cells from the extreme pH conditions in the bulk solution by its attachment to surfaces as shown in detail in Fig. 5b.
In this research, we observed the production of nano-jarosite precipitates at pH 1.1–1.3 during continuous ferrous oxidation by thermoacidophilic archaeon culture. Jarosite formation is favoured at a pH between 1.7 and 2.3 in the temperature range of 60–100°C (Das et al. 1996). Jarosites of biogenic origin are produced at pH values between 2 to 2.5 (Bigham et al. 2010; Wang et al. 2007; Gramp et al. 2008; Egal et al. 2009; Gramp et al. 2009). Despite the low pH, it seems that the stability domains of jarosite at these temperatures (70–75 °C) shift towards a more acidic region. In this acidic region (0.1 M H2SO4), jarosite remains stable and it will dissolve above 0.5 M H2SO4 (Das et al. 1996). XRD analysis suggested that the main precipitated formed is K-jarosite. This could be related to the addition of ammonium and potassium to the medium. However, the formation of ammonium-jarosite only takes place above 160 mM NH +4 (Wang et al. 2007; Gramp et al. 2008), whereas our medium only contained 1.03 mM NH +4 , therefore we consider the formation of ammonium jarosites unlikely
K-jarosite only needs very low K concentrations to be produced (4 mM K+) (Gramp et al. 2008), while our medium contained 8.24 mM K+. Based on the absence of ammonio-jarosite production, it is unlikely that the biomass nitrogen measures were overlapped by nitrogen contained in the precipitates. More importantly, it also dismisses the possibility that the nitrogen source was stripped out from the medium.
The objective of this research was to study the growth of thermoacidophilic cultures at suboptimal pH and substrate conditions to be used in biomineral production. Biomass retention by using activated carbon as carrier material was tested, but this was not successful because the activated carbon disintegrated at the applied conditions. However, it was observed in mesophilic high fluidized-bed reactor a tenfold increase in the iron oxidation rates when jarosite was used as carrier material for biomass instead of activated carbon (Kinnunen and Puhakka 2004). This suggests that jarosite may be used as carrier material in thermoacidophilic reactors to increase the ferrous iron oxidation rates.
The low pH of 1.1 strongly affected the growth rate of the microorganisms, but it did not affect the iron oxidation capacity. Thus, it may be possible to grow the culture at optimal pH conditions to increase cell density and to use these cells for iron oxidation to mediate biomineral formation in continuous processes. In this study, we demonstrated that it is feasible to grow thermoacidophilic microorganisms in continuous bioreactors at pH values and substrate concentrations below what is considered to be optimum values for growth.
References
Astudillo C, Acevedo F (2009) Effect of CO2 air enrichment in the biooxidation of a refractory gold concentrate by Sulfolobus metallicus adapted to high pulp densities. Hydrometallurgy 97(1–2):94–97
Auernik KS, Maezato Y, Blum PH, Kelly RM (2008) The genome sequence of the metal-mobilizing, extremely thermoacidophilic archaeon Metallosphaera sedula provides insights into bioleaching-associated metabolism. Appl Environ Microbiol 74(3):682–692
Becker T, Gorham N, Shiers DW, Watling HR (2011) In situ imaging of Sulfobacillus thermosulfidooxidans on pyrite under conditions of variable pH using tapping mode atomic force microscopy. Process Biochem 46(4):966–976
Bertoldo C, Dock C, Antranikian G (2004) Thermoacidophilic microorganisms and their novel biocatalysts. Eng Life Sci 4(6):521–532
Bigham JM, Jones FS, Ã-zkaya B, Sahinkaya E, Puhakka JA, Tuovinen OH (2010) Characterization of jarosites produced by chemical synthesis over a temperature gradient from 2 to 40 °C. Int J Miner Process 94(3–4):121–128
Bosecker K (1997) Bioleaching: metal solubilization by microorganisms. FEMS Microbiol Rev 20(3–4):591–604
Das GK, Acharya S, Anand S, Das RP (1996) Jarosites: a review. Miner Process Extr Metall Rev 16(3):185–210
De Kock SH, Barnard P, Du Plessis CA (2004) Oxygen and carbon dioxide kinetic challenges for thermophilic mineral bioleaching processes. Biochem Soc Trans 32(2):273–275
Dinkla IJT, Gericke M, Geurkink BK, Hallberg KB (2009) Acidianus brierleyi is the dominant thermoacidophile in a bioleaching community processing chalcopyrite containing concentrates at 70°C. Adv Mater Res 71–73:67–70
Egal M, Casiot C, Morin G, Parmentier M, Bruneel O, Lebrun S, Elbaz-Poulichet F (2009) Kinetic control on the formation of tooeleite, schwertmannite and jarosite by Acidithiobacillus ferrooxidans strains in an As(III)-rich acid mine water. Chem Geol 265(3–4):432–441
Etzel K, Klingl A, Huber H, Rachel R, Schmalz G, Thomm M, Depmeier W (2008) Etching of 111 and 210 synthetic pyrite surfaces by two archaeal strains, Metallosphaera sedula and Sulfolobus metallicus. Hydrometallurgy 94(1–4):116–120
Fuchs T, Huber H, Teiner K, Burggraf S, Stetter KO (1996) Metallosphaera prunae, sp. nov., a novel metal-mobilizing, thermoacidophilic Archaeum, isolated from a uranium mine in Germany. Syst Appl Microbiol 18(4):560–566
Gericke M, Pinches A (1999) Bioleaching of copper sulphide concentrate using extreme thermophilic bacteria. Miner Eng 12(8):893–904
Gericke M, Pinches A, Van Rooyen JV (2001) Bioleaching of a chalcopyrite concentrate using an extremely thermophilic culture. Int J Miner Process 62(1–4):243–255
Gonzalez-Contreras P, Weijma J, Buisman CJN (2009) Arsenic immobilization by biological scorodite crystallization: effect of high ferric concentration and foreign seeds. Adv Mat Res 73:629–632
Gonzalez-Contreras P, Weijma J, Rvd W, Buisman CJN (2010) Biogenic scorodite crystallization by Acidianus sulfidivorans for arsenic removal. Environ Sci Technol 44(2):675–680
Gramp JP, Jones FS, Bigham JM, Tuovinen OH (2008) Monovalent cation concentrations determine the types of Fe(III) hydroxysulfate precipitates formed in bioleach solutions. Hydrometallurgy 94(1–4):29–33
Gramp JP, Wang H, Bigham JM, Sandy Jones F, Tuovinen OH (2009) Biogenic synthesis and reduction of Fe(III)-hydroxysulfates. Geomicrobiol J 26(4):275–280
Hatzinikolaou DG, Kalogeris E, Christakopoulos P, Kekos D, Macris BJ (2001) Comparative growth studies of the extreme thermophile Sulfolobus acidocaldarius in submerged and solidified substrate cultures. World J Microbiol Biotechnol 17(3):229–234
Huber H, Stetter KO (1998) Hyperthermophiles and their possible potential in biotechnology. J Biotechnol 64(1):39–52
Janssen AJH, Lens PNL, Stams AJM, Plugge CM, Sorokin DY, Muyzer G, Dijkman H, Van Zessen E, Luimes P, Buisman CJN (2009) Application of bacteria involved in the biological sulfur cycle for paper mill effluent purification. Sci Total Environ 407(4):1333–1343
Karavaiko GI, Dubinina GA, Kondrateva TF (2006) Lithotrophic microorganisms of the oxidative cycles of sulfur and iron. Microbiology 75(5):512–545
Kinnunen PHM, Puhakka JA (2004) High-rate ferric sulfate generation by a Leptospirillium ferriphilum-dominated biofilm and the role of jarosite in biomass retainment in a fluidized-bed reactor. Biotechnol Bioeng 85(7):697–705
Mikkelsen D, Kappler U, Webb RI, Rasch R, McEwan AG, Sly LI (2007) Visualisation of pyrite leaching by selected thermophilic archaea: nature of microorganism-ore interactions during bioleaching. Hydrometallurgy 88(1–4):143–153
Nemati M, Harrison STL (2000) Comparative study on thermophilic and mesophilic biooxidation of ferrous iron. Miner Eng 13(1):19–24
Nemati M, Lowenadler J, Harrison STL (2000) Particle size effects in bioleaching of pyrite by acidophilic thermophile Sulfolobus metallicus (BC). Appl Microbiol Biotechnol 53(2):173–179
Norris PR (2006) Acidophile diversity in mineral sulfide oxidation. In: Ralings DE, Johnson BD (eds) Biomining. Springer, New York, pp 201–216
Norris PR, Burton NP, Foulis NAM (2000) Acidophiles in bioreactor mineral processing. Extremophiles 4(2):71–76
Plessis CAD, Batty JD, Dew DW (2006) Comercial applications of thermophile bioleaching. In: Ralings DE, Johnson BD (eds) Biomining. Springer, New York, pp 57–80
Plumb JJ, Gibbs B, Stott MB, Robertson WJ, Gibson JAE, Nichols PD, Watling HR, Franzmann PD (2002) Enrichment and characterisation of thermophilic acidophiles for the bioleaching of mineral sulphides. Miner Eng 15(11):787–794
Plumb JJ, Haddad CM, Gibson JAE, Franzmann PD (2007) Acidianus sulfidivorans sp. nov., an extremely acidophilic, thermophilic archaeon isolated from a solfatara on Lihir Island, Papua New Guinea, and emendation of the genus description. Int J Syst Evol Microbiol 57(7):1418–1423
Ralings DE (2006) Relevance of cell physiology and genetic adaptability of biomining microorganisms to industrial processes. In: Ralings DE, Johnson BD (eds) biomining. Springer, New York, pp 177–198
Wang H, Bigham JM, Jones FS, Tuovinen OH (2007) Synthesis and properties of ammoniojarosites prepared with iron-oxidizing acidophilic microorganisms at 22–65 °C. Geochim Cosmochim Acta 71(1):155–164
Witne JY, Phillips CV (2001) Bioleaching of Ok Tedi copper concentrate in oxygen- and carbon dioxide-enriched air. Miner Eng 14(1):25–48
Zeng W, Tan S, Chen M, Qiu G (2011) Detection and analysis of attached microorganisms on the mineral surface during bioleaching of pure chalcopyrite with moderate thermophiles. Hydrometallurgy 106(1–2):46–50
Acknowledgements
This research was financially supported by the Dutch Ministry of Economic Affairs/AgentschapNL through the INNOWator programme, Paques B.V (Balk, The Netherlands), and Nyrstar (Budel-Dorplein, the Netherlands). The authors acknowledge the personal scholarship from CONICYT-Chile to P. Gonzalez. We greatly acknowledge the assistance with SEM–EDX analysis to Jacqueline Donkers from AFSG-WUR Biobased products.
Open Access
This article is distributed under the terms of the Creative Commons Attribution Noncommercial License which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Supplementary Fig. S1
(DOCX 178 kb)
Supplementary Fig. S2
(DOCX 109 kb)
Supplementary Fig. S3
(DOCX 577 kb)
Supplementary Fig. S4
(DOCX 1944 kb)
Rights and permissions
Open Access This is an open access article distributed under the terms of the Creative Commons Attribution Noncommercial License (https://creativecommons.org/licenses/by-nc/2.0), which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
About this article
Cite this article
Gonzalez-Contreras, P., Weijma, J. & Buisman, C.J.N. Kinetics of ferrous iron oxidation by batch and continuous cultures of thermoacidophilic Archaea at extremely low pH of 1.1–1.3. Appl Microbiol Biotechnol 93, 1295–1303 (2012). https://doi.org/10.1007/s00253-011-3460-7
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00253-011-3460-7